Дельта-8 десатураза та її застосування для одержання поліненасичених жирних кислот







Формула / Реферат
1. Виділений полінуклеотид, який відрізняється тим, що включає:
(a) нуклеотидну послідовність, що кодує поліпептид, який має активність дельта-8 десатурази, де поліпептид демонструє як мінімум 80 % ідентичність амінокислот, за даними вирівнювання способом Clustal W, в порівнянні з послідовністю амінокислот, представленою в SEQ ID NO: 47;
(b) нуклеотидну послідовність, що кодує поліпептид, який має активність дельта-8 десатурази, де нуклеотидна послідовність демонструє як мінімум 80 % ідентичність послідовності, за даними вирівнювання способом BLASTN, в порівнянні з нуклеотидною послідовністю, представленою в SEQ ID NO: 92;
(c) нуклеотидну послідовність, що кодує поліпептид, який має активність дельта-8 десатурази, де нуклеотидна послідовність гібридизується в жорстких умовах з нуклеотидною послідовністю, представленою в SEQ ID NO: 92; або
(d) послідовність, комплементарну послідовності (а) нуклеотиду, (b) або (с), де комплементарна і нуклеотидна послідовність складаються з однакової кількості нуклеотидів і є на 100 % комплементарними.
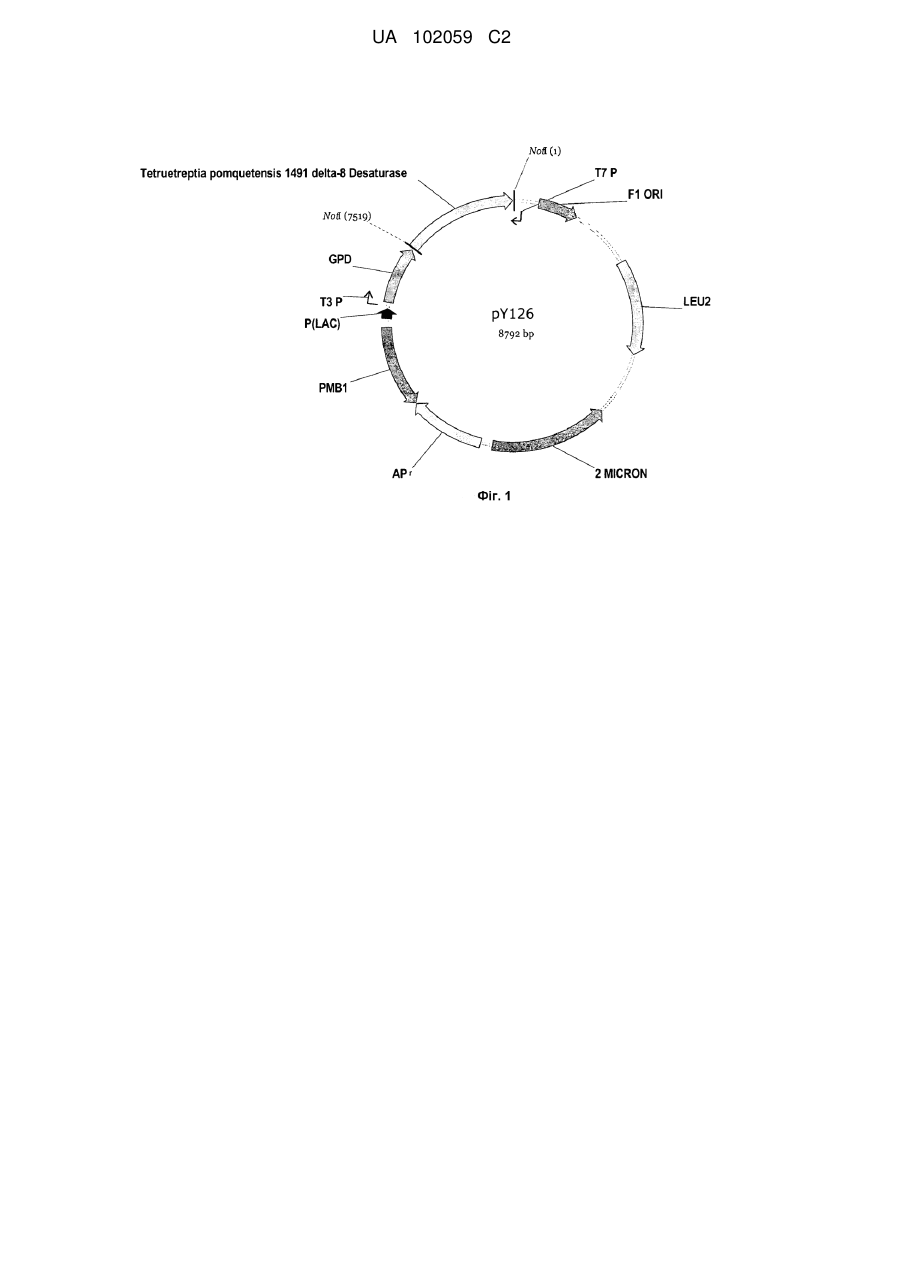
2. Полінуклеотид за п. 1, який відрізняється тим, що нуклеотидна послідовність включає SEQ ID NO: 92.
3. Поліпептид за п. 1, який відрізняється тим, що послідовність амінокислот поліпептиду включає SEQ ID NO: 47.
4. Рекомбінантний конструкт ДНК, який відрізняється тим, що включає полінуклеотид за п. 1 або п. 2, функціонально зв'язаний як мінімум з однією регуляторною послідовністю.
5. Клітина, яка відрізняється тим, що містить в геномі рекомбінантний конструкт ДНК за п. 4 .
6. Рослина олійної культури, яка відрізняється тим, що містить в геномі рекомбінантний конструкт за п. 4.
7. Трансгенне зерно, яке відрізняється тим, що містить в геномі рекомбінантний конструкт за п. 4.
8. Соєва олія, що містить 13,1-17,9 мас. % гексадеканової кислоти, 2,0-3,4 мас. % октадеканової кислоти, 11,8-25,5 мас. % цис-9-октадеканової кислоти, 17,9-27,9 мас. % цис-9,12-октадекадієнової кислоти, 0,3-1,6 мас.% цис-6,9,12-октадекатриєнової кислоти, 6,7-10,1 мас. % цис-9,12,15-октадекатриєнової кислоти, 1,7-9,5 мас. % цис-11,14-ейкозадієнової кислоти, 2,1-8,0 мас. % цис-8,11,14-ейкозатриєнової кислоти, 0,9-3,3 мас. % цис-11,14,17-ейкозатриєнової кислоти, 0,8-4,7 мас. % цис-5,11,14,17-ейкозатетраєнової кислоти, 0,7-11,5 мас. % цис-8,11,14,17-ейкозатетраєнової кислоти,3,3-14,3 мас. % цис-5,8,11,14,17-ейкозапентаєнової кислоти та 0,1-0,4 мас. % цис-7,10,13,16,19-докозапентаєнової кислоти.
9. Соєва олія за п. 8, яка відрізняється тим, що додатково містить до 2,8 мас. % цис-5,11,14-ейкозатриєнової кислоти.
10. Соєва олія за п. 8, яка відрізняється тим, що додатково містить до 10,3 мас. % цис-5,8,11,14-ейкозатриєнової кислоти.
11. Рослини-нащадки, які відрізняються тим, що одержані із зерна за п. 7.
12. Харчовий продукт, який відрізняється тим, що містить соєву олію за п. 8.
13. Кормовий продукт, який відрізняється тим, що містить соєву олію за п. 8.
14. Спосіб одержання трансформованої рослини, який відрізняється тим, що включає трансформацію рослинної клітини за допомогою полінуклеотиду за п. 1 або п. 2 та вирощування рослини з трансформованої рослинної клітини.
15. Трансгенне зерно, яке відрізняється тим, що одержане з рослини, одержаної за способом у відповідності до п. 14.
16. Рослини-нащадки, які відрізняються тим, що одержані з зерна за п. 15.
17. Спосіб одержання поліненасичених жирних кислот з довгим ланцюгом в клітині, яка містить знижений рівень побічних продуктів у вигляді жирних кислот, який відрізняється тим, що вказаний спосіб включає:
(a) трансформацію клітини-хазяїна як мінімум за допомогою одного рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, який кодує як мінімум дві дельта-8 десатурази, визначений у п. 1, функціонально зв'язаний як мінімум з однією регуляторною послідовністю; і
(b) селекцію одержаних трансформованих клітин-хазяїв, які містять знижений рівень побічних продуктів у вигляді жирних кислот, в порівнянні з рівнем таких метаболічних побічних продуктів у вигляді жирних кислот в трансформованій клітині-хазяїні, що містять як мінімум один рекомбінантний конструкт ДНК, який включає виділений полінуклеотид, що кодує одну дельта-8 десатуразу, функціонально зв'язаний з регуляторною послідовністю.
18. Спосіб одержання як мінімум однієї поліненасиченої жирної кислоти в клітині рослини олійної культури, який відрізняється тим, що включає:
(a) трансформацію клітини рослини олійної культури за допомогою першого рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, який кодує як мінімум один поліпептид дельта-8 десатурази, визначений у п. 1, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, і як мінімум одного додаткового рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, де кодуючий поліпептид вибраний з групи, що складається з дельта-4 десатурази, дельта-5 десатурази, дельта-6 десатурази, дельта-8 десатурази, дельта-12 десатурази, дельта-15 десатурази, дельта-17 десатурази, дельта-9 десатурази, дельта-9 елонгази, С14/16 елонгази, С16/18 елонгази, С18/20 елонгази і С20/22 елонгази;
(b) вирощування рослини олійної культури з трансформованої клітини із стадії (а); і
(c) селекцію зерен, одержаних з рослин із стадії (b), що містять змінений рівень поліненасичених жирних кислот в порівнянні з рівнем в зернах, одержаних з нетрансформованої рослини олійної культури.
Текст
Реферат: Винахід належить до виділених фрагментів нуклеїнових кислот і рекомбінантного конструкта, що включають такі фрагменти, які кодують дельта-8 десатуразу, а також до олійної рослини, трансгенного зерна, до способу одержання трансформованої рослини, до способу одержання поліненасичених жирних кислот з довгим ланцюгом (ПНЖК) шляхом застосування вказаної дельта-8 десатурази в рослинах, до харчового та кормового продукту. UA 102059 C2 (12) UA 102059 C2 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Дана заявка претендує на пріоритет Тимчасової Заявки США № 60/853,563, поданої 23 жовтня 2006 р., повний зміст якої включений до даного опису шляхом посилання. ГАЛУЗЬ ВИНАХОДУ Даний винахід відноситься до галузі біотехнології. Більш конкретно, даний винахід стосується полінуклеотидних послідовностей, що кодують дельта-8 десатурази, та застосування таких десатураз для одержання поліненасичених жирних кислот з довгим ланцюгом (ПНЖК). ПЕРЕДУМОВИ ВИНАХОДУ Важливість ПНЖК не підлягає сумніву. Наприклад, деякі ПНЖК є важливими біологічними компонентами здорових клітин і визнані як: "ессенціальні" жирні кислоти, які не можуть бути синтезовані de novo у ссавців і, таким чином, повинні бути одержані з раціоном або одержані подовженням та утворенням ненасичених зв'язків з лінолевої кислоти (ЛК; 18:2 -6) або ліноленової кислоти (AЛК; 18:3 -3); складові плазмових мембран клітин, де вони можуть бути знайдені в таких формах як фосфоліпіди або триацилгліцериди; необхідні для належного розвитку (особливо в дитячому мозку, що розвивається) і для утворення та відновлення тканин; і, прекурсори для декількох біологічно активних ейкозаноїдів, важливих для ссавців (наприклад, простацикліни, ейкозаноїди, лейкотрієни, простагландини). Додатково, високе споживання -3 ПНЖК з довгим ланцюгом здійснює захисний вплив на серцево-судинну систему (Dyerberg et al., Amer. J. Clin. Nutr. 28:958-966 (1975); Dyerberg et al., Lancet. 2(8081):117-119 (1978); Shimokawa, H., World Rev. Nutr. Diet 88:100-108 (2001); von Schacky et al., World Rev. Nutr. Diet 88:90-99 (2001)). В численних інших дослідженнях задокументовано широкий спектр сприятливого впливу на стан здоров'я в результаті введення омега-3 та/або омега-6 ПНЖК проти різноманітних симптомів і захворювань (наприклад, астма, псоріаз, екзема, діабет, рак). На сьогоднішній день, численними зусиллями дослідників різноманітні хазяї, в тому числі рослини, морські водорості, гриби і дріжджі, досліджені як засоби для комерційного продукування ПНЖК. Хоча природна ПНЖК-продукуюча здатність організмів-хазяїв іноді має велике значення для даної методології, генетичне конструювання також довело, що природна здатність деяких хазяїв (навіть, якщо вона від природи обмежена продукуванням жирних кислот ЛК та AЛК) може бути істотною мірою змінена таким чином, щоб приводити до продукування високих рівнів різноманітних омега-3/омега-6 ПНЖК з довгим ланцюгом. Незалежно від того, чи є даний вплив результатом природної здатності або рекомбінантної технології, продукування арахідонової кислоти (АРК; 20:4 -6), ейкозапентаєнової кислоти (ЕПК; 20:5 -3) і докозагексаєнової кислоти (ДГК; 22:6 -3) вимагає експресії шляху дельта-9 елонгази/дельта-8 десатурази (який діє в деяких організмах, таких як види евгленових, і який характеризується продукуванням ейкозадієнової кислоти [ЕДК; 20:2 -6] та/або ейкозатрієнової кислоти [ЕТрК; 20:3 -3], або шляху дельта-6 десатурази/дельта-6 елонгази [який в основному знайдений в морських водоростях, моху, грибках, нематодах та у людини, і який характеризується продукуванням -лінолевої кислоти (ГЛК; 18:3 -6) та/або стеаридонової кислоти (СТК; 18:4 -3) (фіг. 6)]). Дельта-6 елонгаза також відома як C18/20 елонгаза. Ферменти дельта-8 десатурази, ідентифіковані до цього часу, володіють здатністю перетворювати як ЕДК на дигомо--ліноленову кислоту (ДГЛК; 20:3), так і ЕТрК на ейкозатетраєнову кислоту (ЕТК; 20:4) (де АРК та ЕПК послідовно синтезуються з ДГЛК і ЕТК, відповідно, після реакції з дельта-5 десатуразою, тоді як синтез ДГК вимагає додаткової експресії C20/22 елонгази і дельта-4 десатурази). На базі ролі ферментів дельта-8 десатурази в синтезі, наприклад, АРК, ЕПК і ДГК, були здійснені значні зусилля для ідентифікації та характеристики цих ферментів. Більшість зусиль до цього часу були зосереджені на виділенні і характеристиці дельта-8 десатурази з Euglena gracilis; і повідомлялося про декілька варіацій послідовності в межах дельта-8 десатурази Euglena gracilis (див., наприклад, Wallis et al., Arch. Biochem. і Biophys. 365(2):307-316 (1999); публікація PCT WO 2000/34439; патент США 6,825,017; публікація PCT WO 2004/057001). Також, заявки США патентного повіреного Заявників, що розглядаються, за номерами №№ 11/166,003 і 11/166,993, подані 24 червня 2005 р. (номери патентного повіреного BB-1547 і CL3150, відповідно [публікації PCT WO 2006/012325 і WO 2006/012326; обидві опубліковані 2 лютого 2006 р.]) розкривають послідовності амінокислот і нуклеїнових кислот для дельта-8 десатурази Euglena gracilis. В більш пізній публікації PCT WO 2005/103253 (опублікована 22 квітня 2005 р.) розкриті послідовності амінокислот і нуклеїнових кислот для ферменту дельта-8 десатурази з Pavlova salina (див. також Публікацію США № 2005/0273885). Sayanova et al. (FEBS Lett. 580:1946-1952 (2006)) описують виділення і характеристику кДНК земляної амеби Acanthamoeba castellanii, яка при експресії в Arabidopsis кодує дельта-8 десатуразу C20. Також, в заявці патентного 1 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 повіреного Заявників, що розглядається, а саме тимчасовій заявці № 60/795,810, поданій 28 квітня 2006 р. (номер патентного повіреного BB-1566), розкриті послідовності амінокислот і нуклеїнових кислот для ферменту дельта-8 десатурази з Pavlova lutheri (CCMP459). Базуючись на придатності до експресії дельта-8 десатурази в сполученні з дельта-9 елонгазою, також було докладено значних зусиль для ідентифікації та характеристики дельта-9 елонгази з різних джерел. Дельта-9 елонгаза з Isochrysis galbana публічно доступна (описана в GenBank за інвентарним номером AAL37626, а також в PCT публікації WO 02/077213). В заявці патентного повіреного Заявників, а саме тимчасовій заявці США № 60/739,989, поданій 23 листопада 2005 р. (номер патентного повіреного BB-1562), розкрита дельта-9 елонгаза з Eulgena gracilis. Представником Заявників подано цілий ряд патентних заявок, що стосуються продукування ПНЖК в маслянистих дріжджах (тобто, Yarrowia lipolytica), зокрема: PCT публікації WO 2004/101757 і WO 2004/101753 (обидві опубліковані 25 листопада 2004 р.); заявка США № 11/265,761 (подана 2 листопада 2005 р.); заявка США № 11/264,784 (подана 1 листопада 2005 р.); і заявка США № 11/264,737 (подана 1 листопада 2005 р.). Подібним чином, публікація PCT WO 2004/071467 (опублікована 26 серпня, 2004; номер патентного повіреного BB-1538) стосується продукування ПНЖК в рослинах, тоді як PCT публікація WO 2004/071178 (опублікована 26 серпня 2004 р.) стосується анексинових промоторів і їх застосування для експресії трансгенів в рослинах; обидві - заявки представника Заявників, що розглядаються. КОРОТКИЙ ОПИС ВИНАХОДУ Даний винахід стосується виділеного полінуклеотиду, який включає: (a) нуклеотидну послідовність, що кодує поліпептид, який володіє активністю дельта-8 десатурази, де поліпептид демонструє як мінімум 80 % ідентичність амінокислот, за даними вирівнювання способом Clustal V, в порівнянні з послідовністю амінокислот, представленою в SEQ ID NO: 47, SEQ ID NO: NO: 49 або SEQ ID NO: 57; (b) нуклеотидну послідовність, що кодує поліпептид, який володіє активністю дельта-8 десатурази, де нуклеотидна послідовність демонструє як мінімум 80 % ідентичність послідовності, за даними вирівнювання способом BLASTN, в порівнянні з нуклеотидною послідовністю, представленою в SEQ ID NO: 92, SEQ ID NO: 93 або SEQ ID NO: 62; (c) нуклеотидну послідовність, що кодує поліпептид, який володіє активністю дельта-8 десатурази, де нуклеотидна послідовність гібридизується в жорстких умовах з нуклеотидною послідовністю, представленою в SEQ ID NO: 92, SEQ ID NO: 93 або SEQ ID NO: 62; або (d) послідовність, комплементарну нуклеотидній послідовності (a), (b) або (c), де комплементарна і нуклеотидна послідовність складаються з однакової кількості нуклеотидів і є на 100 % комплементарними. В другому варіанті, винахід стосується рекомбінантного конструкту ДНК, що включає будьякий з виділених полінуклеотидів за винаходом, функціонально зв'язаний як мінімум з однією регуляторною послідовністю. У третьому варіанті, винахід стосується клітини, що містить в геномі рекомбінантний конструкт ДНК за винаходом. Такі клітини можуть бути рослинними клітинами або клітинами дріжджів. У четвертому варіанті, винахід стосується способу трансформації клітини, який включає трансформацію клітини за допомогою рекомбінантного конструкта за винаходом або виділеного полінуклеотиду за винаходом, і селекцію клітин, трансформованих за допомогою рекомбінантного конструкта або виділеного полінуклеотиду. У п'ятому варіанті, винахід стосується трансгенного зерна, що містить в геномі рекомбінантний конструкт за винаходом, або трансгенного зерна, одержаного з рослини, одержаної у відповідності до способу за винаходом. Також охоплюється олія або побічні продукти, одержані з таких трансгенних зерен. У шостому варіанті, винахід стосується способу одержання поліненасичених жирних кислот з довгим ланцюгом у рослинній клітині, який включає: (a) трансформацію клітини за допомогою рекомбінантного конструкта за винаходом; і (b) селекцію таких трансформованих клітин, які продукують поліненасичені жирні кислоти з довгим ланцюгом. У сьомому варіанті, винахід стосується способу одержання як мінімум однієї поліненасиченої жирної кислоти у клітині рослини олійної культури, який включає: (а) трансформацію клітини рослини олійної культури за допомогою першого рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, який кодує як мінімум один поліпептид дельта-8 десатурази, функціонально зв'язаний як мінімум з однією 2 UA 102059 C2 5 10 15 20 25 30 35 40 45 регуляторною послідовністю. і як мінімум одного додаткового рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, який кодує поліпептид, вибраний з групи, що складається з дельта-4 десатурази, дельта-5 десатурази, дельта-6 десатурази, дельта-8 десатурази, дельта12 десатурази, дельта-15 десатурази, дельта-17 десатурази, дельта-9 десатурази, дельта-9 елонгази, C14/16 елонгази, C16/18 елонгази, C18/20 елонгази і C20/22 елонгази; (b) регенерацію рослини олійної культури з трансформованої клітини із стадії (a); і (c) селекцію таких зерен, одержаних з рослин стадії (b), що містять змінений рівень поліненасичених жирних кислот в порівнянні з рівнем в зернах, одержаних з нетрансформованої рослини олійної культури. У восьмому варіанті, винахід стосується рослини олійної культури, що містить в геномі рекомбінантний конструкт за винаходом. Придатні рослини олійної культури включають, не обмежуючись ними, сою, види Brassica, соняшник, маїс, бавовну, льон і сафлор. У дев'ятому варіанті, винахід стосується рослини олійної культури, що містить: (a) перший рекомбінантний конструкт ДНК, що включає виділений полінуклеотид, який кодує як мінімум один поліпептид дельта-8 десатурази, функціонально зв'язаний як мінімум з однією регуляторною послідовністю; і (b) як мінімум один додатковий рекомбінантний конструкт ДНК, що включає виділений полінуклеотид, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, який кодує поліпептид, вибраний з групи, що складається з дельта-4 десатурази, дельта-5 десатурази, дельта-6 десатурази, дельта-8 десатурази, дельта-12 десатурази, дельта-15 десатурази, дельта-17 десатурази, дельта-9 десатурази, дельта-9 елонгази, C14/16 елонгази, C16/18 елонгази, C18/20 елонгази і C20/22 елонгази. Також охоплюються трансгенні зерна, одержані з таких рослин олійної культури, а також масла або побічні продукти, одержані з таких трансгенних зерен. Переважним продуктом є лецитин. У десятому варіанті, винахід стосується харчових або кормових продуктів, що містять олію або зерно за винаходом, або харчових або кормових продуктів, що містять інгредієнт, одержаний в результаті обробки зерен. В одинадцятому варіанті, винахід стосується способу одержання поліненасичених жирних кислот з довгим ланцюгом в клітині, що містить знижений рівень побічних продуктів у вигляді жирних кислот, де вказаний спосіб включає: (a) трансформацію клітини-хазяїна за допомогою як мінімум одного рекомбінантного конструкта ДНК, що включає виділений полінуклеотид, що кодує як мінімум дві дельта-8 десатурази, функціонально зв'язаний як мінімум з однією регуляторною послідовністю; і (b) селекцію таких трансформованих клітин-хазяїв, що містять знижений рівень побічних продуктів у вигляді жирних кислот, в порівнянні з рівнем таких метаболічних побічних продуктів у вигляді жирних кислот в трансформованій клітині-хазяїні, що містить як мінімум один рекомбінантний конструкт ДНК, який включає виділений полінуклеотид, що кодує одну дельта-8 десатуразу, функціонально зв'язаний з регуляторною послідовністю. У дванадцятому варіанті, винахід стосується рослин-нащадків, одержаних з рослини, одержаної за способом у відповідності до винаходу, або рослини олійної культури за винаходом. ДЕПОНУВАННЯ БІОЛОГІЧНИХ ЗРАЗКІВ Наступна плазміда депонована а Американській колекції типових культур (ATCC), 10801 бульвар Університету, Манассас, VA, 20110-2209, і має наступне позначення, інвентарний номер і дату депонування (табл. 1). Таблиця 1 Депонування ATCC Плазміда pKR72 50 Інвентарний номер PTA-6019 Дата депонування 28 травня 2004 р. КОРОТКИЙ ОПИС ФІГУР І ПЕРЕЛІКУ ПОСЛІДОВНОСТЕЙ Винахід може бути більш повно зрозумілий з наступного детального опису і доданих фігур та Переліку послідовностей, які складають частину даної заявки. Фіг. 1 представляє вектор експресії дріжджів pY126. Фіг. 2 представляє вектор експресії сої pKR1013. 3 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Фіг. 3 представляє вектор експресії сої pKR1014. Фіг. 4 представляє вектор експресії сої pKR1005. Фіг. 5 представляє ліпідні профілі соматичних соєвих ембріонів, що експресують дельта-8 десатуразу Tetruetreptia pomquetensis CCMP1491 і дельта-9 елонгазу Isochrysis galbana (див. Приклад 10). Фіг. 6 представляє характерний шлях омега-3 та омега-6 жирних кислот, що передбачає перетворення міристинової кислоти на ДГК через різні проміжні сполуки. Фіг. 7A і 7B демонструють вирівнювання методом Clustal V дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 (SEQ ID NO: 57), Eutreptiella sp. CCMP389 (SEQ ID NO: 47), Eutreptiella cf_gymnastica CCMP1594 (SEQ ID NO: 49), Euglena gracilis (SEQ ID NO: 98; інвентарний номер NCBI AAD45877 [GI 5639724]) і Euglena gracilis (SEQ ID NO: 112). Фіг. 8 представляє схематично вектор експресії Yarrowia lipolytica pFBAIn-MOD1. Фіг. 9 представляє схематично конструкт pZKLeuN-29E3. Фіг. 10 представляє ліпідні профілі соматичних соєвих ембріонів, що експресують дельта-9 елонгазу (EgD9e) Tetruetreptia pomquetensis CCMP1491 (TpomD8) і Euglena gracilis для 5 найкращих сортів (див. Приклад 12). Фіг. 11 представляє вектор експресії сої pKR973. Фіг. 12 представляє вектор експресії сої pKR1084. Фіг. 13 представляє вектор експресії сої pKR1123 Фіг. 14 демонструє ліпідні профілі соматичних соєвих ембріонів, що експресують E1594D8 і EgD9e для 5 найкращих сортів. Жирні кислоти ідентифіковані як 16:0 (пальмітат), 18:0 (стеаринова кислота), 18:1 (олеїнова кислота), ЛК, ГЛК, AЛК, ЕДК, ДГЛК, ЕРК і ЕТК; і склад жирних кислот, наведений на Фіг. 14, виражений як відсоток (% мас.) загальної маси жирних кислот. Активність E1594D8 виражена, як відсоток ненасиченості (% ненасич.), обчислений у відповідності до наступної формули: ([продукт]/[субстрат + продукт])*100 Сукупний відсоток ненасиченості для ЕДК і ЕРК показаний як "C 20 % дельта-8 ненасич.". Індивідуальний відсоток ненасиченості дельта-8 омега-6 ("ЕДК % дельта-8 ненасич.") обчислений як: ([ДГЛК]/[ДГЛК + ЕДК])*100 Подібним чином, індивідуальний відсоток ненасиченості дельта-8 омега-3 ("ЕРА % дельта-8 ненасич.") обчислений як: ([ЕТК]/[ЕТК + ЕРА])*100 Співвідношення дельта-8 ненасиченості для омега-6 проти омега-3 субстратів ("співвідношення [ЕДК/ЕРК] % ненасич.") одержане шляхом ділення ЕДК % дельта-8 ненасиченості на ЕРК % дельта-8 ненасиченості. Фіг. 15 представляє вектор експресії сої pKR1122. Фіг. 16 демонструє ліпідні профілі соматичних соєвих ембріонів, що експресують E389D8 і EgD9e для найкращих 5 сортів. Фіг. 17 представляє бінарну експресію Arabidopsis pKR1022R. Фіг. 18 демонструє ліпідні профілі насипного зерна T2 для 22 сортів, де Arabidopsis дикого типу був трансформований pKR1022R (SEQ ID NO: 141). Фіг. 19 демонструє середні профілі жирних кислот (середня величина для 10 ембріонів кожного осрту) соєвих ембріонів, трансформованих за допомогою фрагментів AscI pKR1005 (SEQ ID NO: 90; фіг. 4) і pKR973 (SEQ ID NO: 125; фіг. 11), для 10 сортів, що містять найвищі кількості дельта-8 ненасичених продуктів. Жирні кислоти ідентифіковані як 16:0 (пальмітат), 18:0 (стеаринова кислота), 18:1 (олеїнова кислота), ЛК, ГЛК, AЛК, ЕДК, СКІ, ДГЛК, АРК, ЕРК, ЮНП, ЕТК, ЕПК і ДПК; та їх вміст виражений, як відсоток (% мас.) загальної маси жирних кислот. Жирні кислоти, наведені в переліку як "інші", включають: 18:2 (5,9), СТК, 20:0, 20:1(11), 20:2 (7,11) або 20:2 (8,11), і ДГК. Загальний % (мас.) жирних кислот, що містять подвійний зв'язок дельта-8, виражений як C20 дельта-8 ненасич. (ДГЛК + АРК + ЕТК + ЕПК + ДПК), і активність дельта-8 десатурази виражена як відсоток ненасиченості (C20 % дельта-8 ненасич.), обчислений у відповідності до наступної формули: ([ДГЛК + ЕТК]/[ДГЛК + ЕТК + ЕДК + ЕРК])*100 Фіг. 20 демонструє профілі жирних кислот для 10 індивідуальних зерен T1 з 2 рослин сорту AFS 4882-4-6 (рослини #4882-4-6-1 і #4882-4-6-2), що продемонстрували деякі з найвищих показників загального вмісту продуктів з дельта-8 ненасиченістю. Фіг. 21 демонструє середні профілі жирних кислот (середня величина для 10 ембріонів кожного сорту) соєвих ембріонів, трансформованих за допомогою фрагментів AscI pKR1005 (SEQ ID NO: 90; фіг. 4) і pKR1084 (SEQ ID NO: 129; фіг. 12), для 10 сортів, що містять найвищі 4 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 кількості продуктів з дельта-8 ненасиченістю. Жирні кислоти ідентифіковані як 16:0 (пальмітат), 18:0 (стеаринова кислота), 18:1 (олеїнова кислота), ЛК, ГЛК, AЛК, ЕДК, СКІ, ДГЛК, АРК, ЕРК, ЮНП, ЕТК, ЕПК і ДПК; і склад жирних кислот виражений як відсоток (% мас.) загальної маси жирних кислот. Жирні кислоти, наведені в переліку як "інші", включають: 18:2 (5,9), СТК, 20:0, 20:1(11), 20:2 (7,11) або 20:2 (8,11), і ДГК. Загальний % (мас.) жирних кислот, що містять подвійний зв'язок дельта-8, виражений як дельта-8 C20 ненасич. (ДГЛК + АРК + ЕТК + ЕПК + ДПК), і активність дельта-8 десатурази виражена як відсоток ненасиченості (C20 % дельта-8 ненасич.), обчислений у відповідності до наступної формули: ([ДГЛК + ЕТК]/[ДГЛК + ЕТК + ЕДК + ЕРК])*100 Фіг. 22 демонструє профілі жирних кислот для індивідуальних зерен T1 з 2 рослин сортуAFS 5003-1-8 (рослини #5003-1-8-1 і #5003-1-8-2), що демонструють деякі з найвищих показників загального вмісту продуктів з дельта-8 ненасиченістю. В описі послідовностей наведено коротку характеристику послідовностей, Перелік додається до даного опису. Перелік послідовностей містить однолітерні коди для символів нуклеотидної послідовності, та одно- і трьохлітерні коди для амінокислот, як визначено в стандартах ІЮПАК-МБС (IUPAC-IUB), описаних в Nucleic Acids Research 13:3021-3030 (1985) та в Biochemical Journal 219(2):345-373 (1984). Перелік послідовностей додається до даного опису на компакт-диску. Зміст компакт-диска, що містить Перелік послідовностей, таким чином, включено шляхом посилання у відповідності до 37 C.F.R. §1.52(e). SEQ ID NO: 1-11 представляют нуклеотидні послідовності праймерів D8F1, D8F2, D8F3, D8F4, D8F5, D8F6, D8F7, D8F8, D8F9, D8R1 і D8R2, відповідно. SEQ ID NO: 12 представляє послідовність амінокислот праймерів D8F1 і D8F4. SEQ ID NO: 13 представляє послідовність амінокислот праймерів D8F2, D8F3, D8F5 і D8F6. SEQ ID NO: 14 представляє послідовність амінокислот праймерів D8F7, D8F8 і D8F9. SEQ ID NO: 15 представляє послідовність амінокислот праймерів D8R1 і D8R2. SEQ ID NO: 16 представляє часткову нуклеотидну послідовність дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 із застосуванням комбінації праймерів D8F4/D8R1 (див. Приклад 1). SEQ ID NO: 17 представляє часткову нуклеотидну послідовність дельта-8 десатурази з Eutreptiella sp. CCMP389 із застосуванням комбінації праймерів D8F4/D8R1 (див. Приклад 1). SEQ ID NO: 18 представляє часткову нуклеотидну послідовність дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 із застосуванням комбінації праймерів D8F4/D8R1 (див. Приклад 1). SEQ ID NO: 19 представляє нуклеотидну послідовність олігонуклеотида SMART IV. SEQ ID NO: 20-24 представляють нуклеотидні послідовності праймерів 389D8-3-1, 389D8-32, 389D8-5-1, 389D8-5-2 і 389D8-5-3, відповідно. SEQ ID NO: 25-29 представляють нуклеотидні послідовності праймерів ED8-5-1, ED8-5-2, ED8-5-3, ED8-3-1 і ED8-3-2, відповідно. SEQ ID NO: 30 представляє нуклеотидну послідовність праймера CDSIII/3’ ПЛР. SEQ ID NO: 31 представляє нуклеотидну послідовність адапторного праймера з набору Invitrogen 3’-RACE. SEQ ID NO: 32-36 представляють нуклеотидні послідовності праймерів 1594D8-3-1, 1594D83-2, 1594D8-5-1, 1594D8-5-2 і 1594D8-5-3, відповідно. SEQ ID NO: 37 представляє нуклеотидну послідовність адаптора GenomeWalker (див. також SEQ ID NO: 111). SEQ ID NO: 38 і 39 представляють нуклеотидні послідовності праймерів AP1 і AP2, відповідно. SEQ ID NO: 40 представляє нуклеотидну послідовність pCR2.1-TOPO. SEQ ID NO: 41 представляє нуклеотидну послідовність 5’-ділянки дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 (див. Приклад 2). SEQ ID NO: 42 представляє нуклеотидну послідовність 5’-ділянки дельта-8 десатурази з Eutreptiella sp. CCMP389 (див. Приклад 2). SEQ ID NO: 43 представляє нуклеотидну послідовність 3’-ділянки дельта-8 десатурази з Eutreptiella sp. CCMP389 (див. Приклад 2). SEQ ID NO: 44 представляє нуклеотидну послідовність 3’-ділянки дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 (1594D8-3’A) (див. Приклад 2). SEQ ID NO: 45 представляє нуклеотидну послідовність 3’-ділянки дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 (1594D8-3’B) (див. Приклад 2). 5 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 SEQ ID NO: 46 представляє нуклеотидну послідовність дельта-8 десатурази з Eutreptiella sp. CCMP389 (розмір 1963 пари основ). SEQ ID NO: 47 представляє послідовність амінокислот дельта-8 десатурази з Eutreptiella sp. CCMP389 (кодуюча ділянка SEQ ID NO: 46 і SEQ ID NO: 92). SEQ ID NO: 48 представляє нуклеотидну послідовність дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 (розмір 2063 пари основ). SEQ ID NO: 49 представляє послідовність амінокислот дельта-8 десатурази з Eutreptiella cf_gymnastica CCMP1594 (кодуюча ділянка SEQ ID NO: 48 і SEQ ID NO: 93). SEQ ID NO: 50 представляє нуклеотидну послідовність лінкера TOPO. SEQ ID NO: 51 представляє нуклеотидну послідовність праймера LinkAmp 1. SEQ ID NO: 52 представляє нуклеотидну послідовність праймера LinkAmp 2. SEQ ID NO: 53 представляє нуклеотидну послідовність 5’-ділянки дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 (див. Приклад 3). SEQ ID NO: 54 представляє нуклеотидну послідовність праймера AUAP. SEQ ID NO: 55 представляє нуклеотидну послідовність 3’-ділянки дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 (див. Приклад 3). SEQ ID NO: 56 представляє нуклеотидну послідовність дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 (розмір 2233 пари основ). SEQ ID NO: 57 представляє послідовність амінокислот дельта-8 десатурази з Tetruetreptia pomquetensis CCMP1491 (кодуюча ділянка SEQ ID NO: 56 і SEQ ID NO: 62). SEQ ID NO: 58 і 59 представляють нуклеотидні послідовності TpomNot-5 і TpomNot-3, відповідно. SEQ ID NO: 60 представляє нуклеотидну послідовність праймера T7 SEQ ID NO: 61 представляє нуклеотидну послідовність праймера M13-28Rev SEQ ID NO: 62 представляє нуклеотидну послідовність кодуючої послідовності дельта-8 десатурази Tetruetreptia pomquetensis CCMP1491. SEQ ID NO: 63 представляє нуклеотидну послідовність pLF114-10 SEQ ID NO: 64 представляє нуклеотидну послідовність pY-75. SEQ ID NO: 65 представляє нуклеотидну послідовність pY126 (див. фіг. 1). SEQ ID NO: 66 представляє нуклеотидну послідовність pKR123r. SEQ ID NO: 67 представляє нуклеотидну послідовність pKR1007. SEQ ID NO: 68 представляє нуклеотидну послідовність pKR607. SEQ ID NO: 69 представляє нуклеотидну послідовність pKR1013 (див. фіг. 2). SEQ ID NO: 70 представляє нуклеотидну послідовність кодуючої послідовності дельта-9 елонгази Isochrysis galbana (інвентарний номер NCBI AAL37626 (GI 17226123), локус AAL37626, CDS AF390174; Qi et al., FEBS Lett. 510(3):159-165 (2002)). SEQ ID NO: 71 представляє послідовність частини вставки кДНК з клону Euglena gracilis eeg1c.pk001.n5.f (вставка кДНК на 5’ кінці). SEQ ID NO: 72 представляє послідовність частини вставки кДНК з клону eeg1c.pk001.n5.f (вставка кДНК на 3’ кінці). SEQ ID NO: 73 представляє послідовність клону eeg1c.pk001.n5.f (5’ і 3’ послідовності були вирівняні). SEQ ID NO: 74 представляє кодуючу послідовність дельта-9 елонгази Euglena gracilis з кДНК в клоні eeg1c.pk001.n5.f. SEQ ID NO: 75 представляє послідовність амінокислот дельта-9 елонгази Euglena gracilis з клону eeg1c.pk001.n5.f (кодуюча ділянка SEQ ID NO: 74). SEQ ID NO: 76 представляє послідовність амінокислот ферменту подовження ПНЖК з довгим ланцюгом (дельта-9 елонгаза) з Isochrysis galbana (інвентарний номер NCBI AAL37626 (GI 17226123), локус AAL37626, CDS AF390174) (позначений "IgD9e"). SEQ ID NO: 77 і 78 представляють нуклеотидні послідовності олігонуклеотидних праймерів oEugEL1-1 і oEugEL1-2, відповідно. SEQ ID NO: 79 представляє нуклеотидну послідовність pKR906. SEQ ID NO: 80 представляє нуклеотидну послідовність pKR72 (інвентарний номер ATCC PTA-6019). SEQ ID NO: 81 представляє нуклеотидну послідовність pK912. SEQ ID NO: 82 представляє нуклеотидну послідовність pKR1014 (див. фіг. 3). SEQ ID NO: 83 представляє нуклеотидну послідовність pKR271. SEQ ID NO: 84 представляє нуклеотидну послідовність pKR226. SEQ ID NO: 85 представляє нуклеотидну послідовність pKR886r. 6 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 SEQ ID NO: 86 і 87 представляють нуклеотидну послідовність олігонуклеотидних праймерів oCon1 і oCon2, відповідно. SEQ ID NO: 88 представляє нуклеотидну послідовність pKR179. SEQ ID NO: 89 представляє нуклеотидну послідовність pKR1002. SEQ ID NO: 90 представляє нуклеотидну послідовність pKR1005 (див. фіг. 4). SEQ ID NO: 91 представляє нуклеотидну послідовність універсального праймера M13F. SEQ ID NO: 92 представляє нуклеотидну послідовність кодуючої послідовності дельта-8 десатурази Eutreptiella sp. CCMP389 SEQ ID NO: 93 представляє нуклеотидну послідовність кодуючої послідовності дельта-8 десатурази Eutreptiella cf_gymnastica CCMP1594. SEQ ID NO: 94 представляє нуклеотидну послідовність вектора експресії Yarrowia lipolytica pFBAIn-MOD1. SEQ ID NO: 95 представляє нуклеотидну послідовність вектора експресії Yarrowia lipolytica pFBAIn-389D8. SEQ ID NO: 96 представляє нуклеотидну послідовність вектора експресії Yarrowia lipolytica pFBAIn-1594D8. SEQ ID NO: 97 представляє нуклеотидну послідовність вектора експресії Yarrowia lipolytica pFBAIn-1491D8. SEQ ID NO: 98 представляє послідовність амінокислот гена дельта-8 десатурази жирних кислот Euglena gracilis (інвентарний номер NCBI AAD45877 [GI 5639724]). SEQ ID NO: 98 представляє послідовність амінокислот, кодовану нуклеотидом 14-1273, інвентарний номер NCBI AF139720 (GI 5639723). Показано, що дана дельта-8 десатураза жирних кислот є нефункціональною. SEQ ID NO: 99 і 100 представляють нуклеотидні послідовності праймерів 389D8-F і 389D8-R, відповідно. SEQ ID NO: 101 і 102 представляють нуклеотидні послідовності праймерів 1491D8-F і 1491D8-R, відповідно. SEQ ID NO: 103 і 104 представляють нуклеотидні послідовності праймерів 1594D8-F і 1594D8-R, відповідно. SEQ ID NO: 105 складає 5" ПЛР праймер, який використовують в Прикладі 1. SEQ ID NO: 106 представляє нуклеотидну послідовність плазміди pZKLeuN-29E3 (див. фіг. 9). SEQ ID NO: 107 представляє нуклеотидну послідовність синтетичної дельта-9 елонгази (вперше виділена з Euglena gracilis - див. SEQ ID NO: 74), оптимізованої за кодонами для Yarrowia lipolytica; див. також патентну заявку США № 60/739,989, подану 23 листопада 2005 р. ([номер патентного повіреного BB-1562], позначена "EgD9E" або "EgD9S"). SEQ ID NO: 108 представляє нуклеотидну послідовність послідовності LoxP з Escherichia coli. SEQ ID NO: 109 представляє нуклеотидну послідовність синтетичної C 16/18 елонгази (вперше виділена з M. alpina), оптимізованої за кодонами для Yarrowia lipolytica; див. також патентну заявку США № 11/253882, подану 19 жовтня 2005 р. SEQ ID NO: 110 представляє нуклеотидну послідовність синтетичної дельта-9 елонгази (вперше виділена з Isochrysis galbana), оптимізованої за кодонами для Yarrowia lipolytica (позначена "IgD9eS"). SEQ ID NO: 111 представляє нуклеотидну послідовність адаптора GenomeWalker (див. також SEQ ID NO: 37). SEQ ID NO: 112 представляє послідовність амінокислот дельта-8 десатурази Euglena gracilis (SEQ ID NO: 2 публікації США № 20050287652). SEQ ID NO: 113 представляє нуклеотидну послідовність pKR132. SEQ ID NO: 114 представляє нуклеотидну послідовність pKR953. SEQ ID NO: 115 представляє нуклеотидну послідовність pKR287. SEQ ID NO: 116 представляє нуклеотидну послідовність дельта-5 десатурази Mortierella alpina (описана в патенті США 6,075,183). SEQ ID NO: 117 представляє нуклеотидну послідовність pKR277. SEQ ID NO: 118 представляє нуклеотидну послідовність pKR952. SEQ ID NO: 119 представляє нуклеотидну послідовність pKR457. SEQ ID NO: 120 представляє нуклеотидну послідовність модифікованої Kti-NotI-Kti3’Salb3" касети. 7 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 SEQ ID NO: 121 представляє нуклеотидну послідовність послідовності кодону касети дельти-8 Pavlova lutheri, описаної в тимчасовій заявці США № 60/795,810 і патентній заявці США № 11/737,772. SEQ ID NO: 122 представляє нуклеотидну послідовність олігонуклеотидного праймера PvDES5’Not-1. SEQ ID NO: 123 представляє нуклеотидну послідовність олігонуклеотидного праймера PvDES3’Not-1. SEQ ID NO: 124 представляє нуклеотидну послідовність pKR970. SEQ ID NO: 125 представляє нуклеотидну послідовність pKR973. SEQ ID NO: 126 представляє нуклеотидну послідовність pKS129. SEQ ID NO: 127 представляє нуклеотидну послідовність pKR606. SEQ ID NO: 128 представляє нуклеотидну послідовність pKR804. SEQ ID NO: 129 представляє нуклеотидну послідовність pKR1084. SEQ ID NO: 130 представляє нуклеотидну послідовність pKR908. SEQ ID NO: 131 представляє нуклеотидну послідовність pKR1118 SEQ ID NO: 132 представляє нуклеотидну послідовність pKR1120. SEQ ID NO: 133 представляє нуклеотидну послідовність pKR1123 SEQ ID NO: 134 представляє нуклеотидну послідовність pKR1117. SEQ ID NO: 135 представляє нуклеотидну послідовність pKR1119. SEQ ID NO: 136 представляє нуклеотидну послідовність pKR1122. SEQ ID NO: 137 представляє нуклеотидну послідовність pKR393. SEQ ID NO: 138 представляє нуклеотидну послідовність pKR407. SEQ ID NO: 139 представляє нуклеотидну послідовність pKR1018. SEQ ID NO: 140 представляє нуклеотидну послідовність pKR1020R. SEQ ID NO: 141 представляє нуклеотидну послідовність pKR1022R. ДЕТАЛЬНИЙ ОПИС ВИНАХОДУ Розкриття кожного посилання, процитованого в даному описі, таким чином, включене шляхом посилання у всій його повноті. В даному описі і в доданій формулі винаходу (текст англійською мовою), форми однини "a", "an" і "the" включають посилання на множину, якщо з контексту не є очевидним інше. Таким чином, наприклад, посилання на "рослину" включає множину таких рослин, посилання на "клітину" включає одну або більше клітин та їх еквівалентів, відомих спеціалістам в даній галузі, і т. д. Даний винахід стосується ферментів дельта-8 десатурази і нуклеїнових кислот для їх кодування, виділених з Tetruetreptia pomquetensis CCMP1491, Eutreptiella sp. CCMP389 і Eutreptiella cf_gymnastica CCMP1594 дельта-8. Вони є придатними, серед іншого, для маніпулювання біохімічними шляхами з метою продукування корисних для здоров'я ПНЖК. Таким чином, об'єкт винаходу має численні застосування. ПНЖК або їх похідні, одержані за методологією, розкритою в даному описі, можна застосовувати як замінники харчових продуктів або харчові добавки, особливо в рецептурах дитячого харчування, для пацієнтів, що одержують харчування внутрішньовенно, або для профілактики чи лікування недоїдання. Альтернативно, очищені ПНЖК (або їх похідні) можуть бути введені в кухарські масла, жири або маргарини, таким чином, що при звичайному застосуванні споживач буде одержувати бажану кількість для доповнення раціону. ПНЖК також можуть бути введені в рецептури дитячого харчування, харчові добавки або інші харчові продукти, і можуть знайти застосування як протизапальні або холестерин-знижуючи засоби. Необов'язково, композиції можуть бути використані для фармацевтичного застосування (в медицині або ветеринарії). В даному випадку, ПНЖК загалом вводять перорально, але їх можна вводити будь-яким способом, при якому вони можуть бути успішно засвоєні, наприклад, парентерально (наприклад, підшкірно, внутрішньом'язово або внутрішньовенно), ректально, інтравагінально або місцево (наприклад, як мазь для шкіри або лосьйон). Додаткове введення людині або тварині ПНЖК, продукованих рекомбінантними засобами, може приводити до збільшених рівнів доданих ПНЖК, а також їх метаболічних нащадків. Наприклад, лікування ЕПК може приводити не тільки до збільшення рівнів ЕПК, але й метаболітів ЕПК, таких як ейкозаноїди (тобто, простагландини, лейкотрієни, тромбоксани). Складні регуляторні механізми можуть зробити бажаним поєднання різних ПНЖК, або додавання різних кон'югатів ПНЖК для попередження, контролю або подолання таких механізмів, щоб досягти бажаних рівнів конкретних ПНЖК у особи. В контексті даного опису застосовується цілий ряд термінів і скорочень. Запропоновані наступні визначення. 8 UA 102059 C2 5 10 15 20 25 "Відкрити рамка зчитування", скорочено ВРЗ. "Полімеразна ланцюгова реакція", скорочено ПЛР. "Американська колекція типових культур", скорочено ATCC. "Поліненасичена(і) жирна(і) кислота(и) ", скорочено ПНЖК. "Триацилгліцериди", скорочено ТАГ. Термін "жирні кислоти" позначає аліфатичні кислоти (алканові кислоти) з довгим ланцюгом варіюючої довжини, від приблизно C12 до C22 (хоча відомі кислоти як з довшим, так і з коротшим ланцюгом). Домінуючі значення довжини ланцюга знаходяться між C 16 і C22. Додаткові деталі, що стосуються диференціації поміж "насиченими жирними кислотами" проти "ненасичених жирних кислот", "мононененасичених жирних кислот" проти "поліненасичених жирних кислот" (або "ПНЖК"), та "омега-6 жирних кислот" (-6 або n-6) проти "омега-3 жирних кислот" (-3 або n-3) запропоновані в PCT публікації WO 2004/101757. Жирні кислоти описані в даному описі простою системою зображення знаками "X:Y", де X представляє кількість атомів вуглецю (C) в конкретній жирній кислоті, і Y представляє кількість подвійних зв'язків. Цифра після позначення жирної кислоти вказує на положення подвійного зв'язку, рахуючи з карбоксильного кінця жирної кислоти з афіксом "c" для цис-конфігурації подвійного зв'язку (наприклад, пальмітинова кислота [16:0], стеаринова кислота [18:0], олеїнова кислота [18:1, 9c], петроселінова кислота [18:1, 6c], ЛК [18:2, 9c, 12c], ГЛК [18:3, 6c, 9c, 12c] і AЛК [18:3, 9c, 12c, 15c]). Якщо не вказано інше, 18:1, 18:2 і 18:3 позначають олеїнову кислоту, ЛК і AЛК жирні кислоти, відповідно. Якщо конкретно не вказано інше, передбачається, що подвійні зв'язки мають цис конфігурацію. Наприклад, передбачається, що подвійні зв'язки в положенні 18:2 (9,12) знаходяться в цис конфігурації. Номенклатура, використана для характеристики ПНЖК в даному описі, продемонстрована в табл. 2 нижче. У стовпчику під назвою "скорочені позначення" застосовується довідкова омегасистема для позначення кількості атомів вуглецю, кількості подвійних зв'язків і положення подвійного зв'язка, найближчого до омега атому вуглецю, рахуючи від омега атома вуглецю (який має номер 1 для даної мети). Решта таблиці підсумовує традиційні назви омега-3 і омега6 жирних кислот та їх прекурсорів, скорочення, які будуть застосовуватися в специфікації далі, і хімічні назви для кожної сполуки. 30 9 UA 102059 C2 Таблиця 2 Номенклатура поліненасичених жирних кислот і прекурсорів Традиційна назва Міристинова Пальмітинова Пальмітолеїнова Стеаринова Олеїнова Лінолева гама-Ліноленова Ейкозадієнова дигомо-гама-Ліноленова Скіадонова Скорочення ПК ЛК ГЛК ЕДК ДГЛК СКІ Арахідонова АРК альфа-Ліноленова AЛК Стеаридонова СТК Ейкозатрієнова ЕТрК або ЕРК Ейкозатетраєнова Юніперова 20 25 ДПК Докозагексаєнова 15 ЕПК Докозапентаєнова 10 ЮНП Ейкозапентаєнова 5 ЕТК ДГК Хімічна назва Скорочене позначення тетрадеканова 14:0 гексадеканова 16:0 9-гексадеценова 16:1 октадеканова 18:0 цис-9-октадеценова 18:1 цис-9,12-октадекадієнова 18:2 -6 цис-6,9,12-октадекатрієнова 18:3 -6 цис-11,14-ейкозадієнова 20:2 -6 цис-8,11,14-ейкозатрієнова 20:3 -6 цис-5,11,14-ейкозатрієнова 20:3b -6 цис-5,8,11,1420:4 -6 ейкозатетраєнова цис-9,12,15-октадекатрієнова 18:3 -3 цис-6,9,12,1518:4 -3 октадекатетраєнова цис-11,14,17-ейкозатрієнова 20:3 -3 цис-8,11,14,1720:4 -3 ейкозатетраєнова цис-5,11,14,17-ейкозатрієнова 20:4b -3 цис-5,8,11,14,1720:5 -3 ейкозапентаєнова цис-7,10,13,16,1922:5 -3 докозапентаєнова цис-4,7,10,13,16,1922:6 -3 докозагексаєнова Метаболічний шлях або біосинтетичний шлях в біохімічному значенні, може бути розцінений як серія каталізованих ферментами хімічних реакцій, що відбуваються в межах клітини для досягнення утворення метаболічного продукту, який використовується або зберігається в клітині, або ініціації іншого метаболічного шляху (далі має назву стадії генерації потоку). Багато з цих шляхів детально опрацьовані і включають постадійну модифікацію початкової субстанції для утворення продукту, що має точну цільову хімічну структуру. Термін "біосинтетичний шлях ПНЖК" позначає метаболічний процес, в ході якого олеїнова кислота перетворюється на ЛК, ЕДК, ГЛК, ДГЛК, АРК, AЛК, СТК, ЕТрК, ЕТК, ЕПК, ДПК і ДГК. Даний процес добре описаний в літературі (наприклад, див. PCT публікацію WO 2006/052870). Якщо коротко, даний спосіб включає подовження вуглецевого ланцюга шляхом додавання вуглецевих атомів, і забезпечення ненасиченості молекули через утворення додаткових подвійних зв'язків за допомогою ряду спеціальних ферментів подовження та утворення ненасичених зв'язків (тобто, "ферментів біосинтетичного шляху ПНЖК"), присутніх в мембрані ендоплазматичного ретикулума. Більш конкретно, "фермент біосинтетичного шляху ПНЖК" позначає будь-який з наступних ферментів (і гени, які кодують вказані ферменти), пов'язаних з біосинтезом ПНЖК, зокрема: дельта-4 десатураза, дельта-5 десатураза, дельта-6 десатураза, дельта-12 десатураза, дельта-15 десатураза, дельта-17 десатураза, дельта-9 десатураза, дельта-8 десатураза, дельта-9 елонгаза, C14/16 елонгаза, C16/18 елонгаза, C18/20 елонгаза та/або C20/22 елонгаза. Термін "біосинтетичний шлях омега-3/омега-6 жирних кислот" позначає набір генів, які при експресії за відповідних умов кодують ферменти, які каталізують продукування омега-3 та/або омега-6 жирних кислот. Звичайно гени, залучені до біосинтетичного шляху омега-3/омега-6 жирних кислот, кодують ферменти біосинтетичного шляху ПНЖК. Характерний шлях проілюстрований на фіг. 6 і передбачає перетворення міристинової кислоти на ДГК через різні проміжні ланки, що демонструє, як омега-3 та/або омега-6 жирні кислоти можуть бути продуковані з розповсюдженого джерела. Шлях природним чином розділений на дві частини, де одна частина буде генерувати омега-3 жирні кислоти, а інша частина - омега-6 жирні кислоти. 10 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Термін "функціональний" в даному описі в контексті біосинтетичного шляху омега-3/омега-6 жирних кислот означає, що деякі (або всі) гени в шляху експресують активні ферменти, що приводить до каталізу або перетворення субстрату in vivo. Слід розуміти, що "біосинтетичний шлях омега-3/омега-6 жирних кислот" або "функціональний біосинтетичний шлях омега-3/омега6 жирних кислот" не означає, що всі гени ферментів біосинтетичного шляху ПНЖК є необхідними, оскільки цілий ряд продуктів у вигляді жирних кислот буде вимагати експресії тільки підмножини генів даного шляху. Термін "шлях дельта-9 елонгази/дельта-8 десатурази" позначає біосинтетичний шлях для продукування ПНЖК з довгим ланцюгом. Даний шлях включає як мінімум дельта-9 елонгазу і дельта-8 десатуразу, таким чином, дозволяючи біосинтез ДГЛК та/або ЕТК з ЛК і AЛК, відповідно. При експресії іншої десатурази і елонгази також можуть бути синтезовані АРК, ЕПК, ДПК і ДГК. Даний шлях може бути переважним в деяких варіантах, оскільки виключає біосинтез ГЛК та/або СТК. Термін "проміжна жирна кислота" позначає будь-яку жирну кислоту, яка продукується в метаболічному шляху жирних кислот, яка в даному шляху може бути додатково перетворена на передбачену кінцеву жирну кислоту під дією інших ферментів метаболічного шляху. Наприклад, якщо продукується ЕПК із застосуванням шляху дельта-9 елонгази/дельта-8 десатурази, можуть бути продуковані ЕДК, ЕТрК, ДГЛК, ЕТК і АРК, які вважаються "проміжними жирними кислотами", оскільки ці жирні кислоти можуть бути додатково перетворені на ЕПК під дією інших ферментів метаболічного шляху. Термін "побічний продукт у вигляді жирної кислоти" позначає будь-яку жирну кислоту, продуковану в метаболічному шляху жирних кислот, яка не є ані передбаченою кінцевою жирною кислотою шляху, ані "проміжною жирною кислотою" шляху. Наприклад, якщо продукується ЕПК із застосуванням шляху дельта-9 елонгази/дельта-8 десатурази, скіадонова кислота (СКІ) та юніперова кислота (ЮНП) також можуть бути продуковані під дією дельта-5 десатурази на ЕДК або ЕТрК, відповідно. Вони розглядаються як "побічні продукти у вигляді жирної кислоти", оскільки ні одна з них не може бути додатково трансформована на ЕПК під дією інших ферментів метаболічного шляху. Терміни "триацилгліцерид", "олія" і "ТАГ" позначають нейтральні ліпіди, що складаються з трьох залишків жирних кислот, приєднаних естерним зв'язком до молекули гліцерину (і такі терміни будуть застосовуватися рівнозначним чином в даному описі). Такі олії (масла) можуть містити ПНЖК з довгим ланцюгом, а також насичені і ненасичені жирні кислоти з коротшим ланцюгом і насичені жирні кислоти з довшим ланцюгом. Таким чином, "біосинтез олії" загалом позначає синтез ТАГ в клітині. "Відсоток (%) ПНЖК в загальній кількості ліпідних і масляних фракцій" позначає відсоток ПНЖК відносно загального вмісту жирних кислот в таких фракціях. Терміни "загальна ліпідна фракція" або "ліпідна фракція" обидва позначають сумарну кількість всіх ліпідів (тобто, нейтральних і полярних) в межах маслянистого організму, тобто зокрема ті ліпіди, які розташовані у фосфатидилхоліновій (ФХ) фракції, фосфатидилетаноламіновій (ФЕ) фракції, а також триацилгліцеридній (ТАГ або олійній) фракції. Таким чином, терміни "ліпід" і "олія" застосовуються в описі взаємозамінним чином. Терміни "ефективність перетворення" і "відсоток перетворення субстрату" позначають ефективність, з якою конкретний фермент (наприклад, десатураза) може перетворити субстрат на продукт. Ефективність перетворення вимірюють у відповідності до наступної формули: ([продукт]/[субстрат + продукт])*100, де "продукт" включає безпосередній продукт і всі продукти в шляху, що утворюються з нього. "Десатураза" являє собою поліпептид, який може утворювати ненасиченість, тобто, вводити подвійний зв'язок в одну або більше жирних кислот для утворення цільової жирної кислоти або прекурсора. Незважаючи на застосування в описі довідкової омега-системи для позначення конкретних жирних кислот, більш традиційним є позначення активності десатурази шляхом рахування від карбоксильного кінця субстрату із застосуванням системи дельта. Особливий інтерес в даному описі представляють дельта-8 десатурази, які утворюють подвійний зв'язок в жирній кислоті між восьмим і дев'ятим атомами вуглецю, пронумерованими від кінцевого атома на карбоксильному кінці молекули, і яка може, наприклад, каталізувати перетворення ЕДК до ДГЛК та/або ЕТрК на ЕТК. Інша придатна десатураза жирних кислот включає, наприклад: (1) дельта-5 десатуразу, яка каталізує перетворення ДГЛК до АРК та/або ЕТК до ЕПК; (2) дельта-6 десатуразу, яка каталізує перетворення ЛК до ГЛК та/або AЛК до СТК; (3) дельта-4 десатуразу, яка каталізує перетворення ДПК до ДГК; (4) дельта-12 десатуразу, яка каталізує перетворення олеїнової кислоти до ЛК; (5) дельта-15 десатуразу, яка каталізує перетворення ЛК до AЛК та/або ГЛК до СТК; (6) дельта-17 десатуразу, яка каталізує перетворення АРК до ЕПК та/або 11 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 ДГЛК до ЕТК; і (7) дельта-9 десатуразу, яка каталізує перетворення пальмітинової кислоти до пальмітолеїнової кислоти (16:1) та/або стеаринової кислоти до олеїнової кислоти (18:1). В даній галузі, дельта-15 і дельта-17 десатурази також іноді позначають як "омега-3 десатурази", "w-3 десатурази" та/або "-3 десатурази", на базі їх здатності перетворювати омега-6 жирні кислоти на їх омега-3 двійники (наприклад, перетворення ЛК на AЛК і АРК на ЕПК, відповідно). У деяких варіантах найбільш бажаним є емпіричне визначення специфічності конкретної десатурази жирних кислот шляхом трансформації придатного хазяїна за допомогою гена десатурази жирних кислот і визначення його впливу на профіль жирних кислот хазяїна. Для цілей даного опису, термін "TpomD8" позначає фермент дельта-8 десатуразу (SEQ ID NO: 57), виділений з Tetruetreptia pomquetensis CCMP1491, кодований SEQ ID NO: 62 в даному винаході. Термін "E389D8" позначає фермент дельта-8 десатуразу (SEQ ID NO: 47), виділений з Eutreptiella sp. CCMP389, кодований SEQ ID NO: 92 в даному винаході. Також, термін "E1594D8" позначає фермент дельта-8 десатуразу (SEQ ID NO: 49), виділений з Eutreptiella cf_gymnastica CCMP1594, кодований SEQ ID NO: 93 в даному винаході. Подібним чином, термін "EgD8" позначає фермент дельта-8 десатуразу (SEQ ID NO: 112), виділений з Euglena gracilis. EgD8 є на 100 % ідентичним і функціонально рівноцінним "Eg5", як описано в публікаціях PCT WO 2006/012325 і WO 2006/012326 (SEQ ID NO: 2 публікації США № 20050287652-A1). Термін "система елонгази" позначає набір чотирьох ферментів, які відповідають за подовження вуглецевого ланцюга жирних кислот для утворення жирної кислоти, ланцюг якої на два атоми вуглецю довший за ланцюг субстрату жирної кислоти, на якому система елонгази діє. Більш конкретно, процес подовження відбувається в сполученні з синтетазою жирних кислот, де КоА представляє носій ацилу (Lassner et al., Plant Cell 8:281-292 (1996)). На першій стадії, що, як виявилося, є специфічною для субстрату, а також обмежуючою швидкість, малоніл-КоА конденсується з ацил-КоА з довгим ланцюгом з утворенням діоксиду вуглецю (CO 2) і кетоацил-КоА (де ацильний фрагмент подовжений двома атомами вуглецю). Подальші реакції включають відновлення до -гідроксиацил-КоА, дегідратацію до єноїл-КоА і друге відновлення з утворенням подовженого ацил-КоА. Прикладами реакцій, каталізованих системами елонгази, є перетворення ГЛК до ДГЛК, СТК до ЕТК, ЛК до ЕДК, AЛК до ЕТрК і ЕПК до ДПК. Для цілей даного опису, фермент, що каталізує першу реакцію конденсації (тобто, перетворення малоніл-КоА і ацил-CvoA з довгим ланцюгом на -кетоацил-КоА), буде позначений загалом як "елонгаза". Загалом, селективність елонгаз по відношенню до субстрату є досить широкою, але вона підрозділяється як за довжиною ланцюга, так і за ступенем ненасиченості. Відповідно, елонгази можуть володіти різною специфічністю. Наприклад, C 14/16 елонгаза буде використовувати субстрат C14 (наприклад, міристинову кислоту), C16/18 елонгаза буде використовувати субстрат C16 (наприклад, пальмітат), C18/20 елонгаза буде використовувати субстрат C18 (наприклад, ГЛК, СТК), та C20/22 елонгаза буде використовувати субстрат C20 (наприклад, ЕПК). Подібним чином, "дельта-9 елонгаза" може бути здатною каталізувати перетворення ЛК до ЕДК та/або AЛК до ЕТрК. Важливо відзначити, що деякі елонгази володіють широкою специфічністю, і, таким чином, єдиний фермент може бути здатний до каталізу декількох реакцій елонгази. Таким чином, наприклад, дельта-9 елонгаза може також діяти як C16/18 елонгаза, C18/20 елонгаза та/або C20/22 елонгаза і може володіти альтернативною, але не переважною специфічністю по відношенню до дельта-5 і дельта-6 жирних кислот, таких як ЕПК та/або ГЛК, відповідно. Для цілей даного опису, термін "IgD9e" позначає дельта-9 елонгазу (SEQ ID NO: 76; інвентарний номер NCBI AAL37626 [GI 17226123], локус AAL37626, CDS AF390174; інвентарний номер GenBank № AF390174), виділену з Isochrysis galbana, кодовану SEQ ID NO: 70. На протилежність цьому, термін "IgD9eS" позначає синтетичну (оптимізовану за кодонами) дельта9 елонгазу (SEQ ID NO: 110), що походить від послідовності ДНК дельта-9 елонгази Isochrysis galbana (SEQ ID NO: 70), яка може застосовуватися для експресії в Yarrowia lipolytica. Подібним чином, для цілей даного опису термін "EgD9e" позначає дельта-9 елонгазу (SEQ ID NO: 75), виділену з Euglena gracilis, кодовану SEQ ID NO: 74 (див. Приклад 11 в даному описі). В даному описі, "нуклеїнова кислота" позначає полінуклеотид і включає одно- або двохланцюговий полімер, що складається з дезоксирибонуклеотидних або рибонуклеотидних основ. Нуклеїнові кислоти також можуть включати фрагменти і модифіковані нуклеотиди. Таким чином, терміни "полінуклеотид", "послідовність нуклеїнової кислоти", "нуклеотидна послідовність" або "фрагмент нуклеїнової кислоти" застосовують рівнозначним чином і представляють полімер РНК або ДНК, одно- або двохланцюговий, що необов'язково містить синтетичні, неприродні або модифіковані нуклеотидні основи. Нуклеотиди (звичайно знайдені в 12 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 формі 5’-монофосфату) скорочено позначені однією літерою, як вказано нижче: "A" для аденілату або дезоксиаденілату (для РНК або ДНК, відповідно), "C" для цитидилату або дезоксицитидилату, "G" для гуанілату або дезоксигуанілату, "U" для уридилату, "T" для дезокситимідилату, "R" для пуринів (А бо G), "Y" для піримідинів (C або T), "K" для G або T, "H" для A або C або T, "I" для інозину і "N" - для будь-якого нуклеотиду. Термін "субфрагмент, що є функціонально рівноцінним" і "функціонально рівноцінний субфрагмент" застосовують в даному описі рівнозначним чином. Ці терміни позначають частину або субпослідовність виділеного фрагмента нуклеїнової кислоти, який зберігає здатність змінювати експресію гена або продукувати певний фенотип, незалежно від того, чи кодує фрагмент або субфрагмент активний фермент. Наприклад, фрагмент або субфрагмент може застосовуватися в дизайні химерних генів для продукування бажаного фенотипу в трансформованій рослині. Химерні гени можуть бути розроблені для застосування в пригніченні шляхом зв'язування фрагменту або субфрагменту нуклеїнової кислоти, незалежно від того, чи кодує він активний фермент, в смисловій або антисмисловій орієнтації по відношенню до послідовності рослинного промотору. Термін "консервативний домен" або "мотив" позначає набір амінокислот, консервативних для конкретних положень вздовж вирівняної послідовності еволюційно споріднених білків. Хоча амінокислоти в інших положеннях можуть варіювати поміж гомологічними білками, високою мірою консервативні амінокислоти в конкретних положеннях вказують на амінокислоти, що є істотними для структури, стабільності або активності білка. Оскільки вони ідентифіковані високим ступенем консервативності у вирівняних послідовностях родини гомологічних білків, вони можуть бути використані як ідентифікатори або "підписи" для визначення приналежності білка з вперше визначеною послідовністю до попередньо ідентифікованої родини білків. Терміни "гомологічний", "гомологія", "істотною мірою подібний" та "істотною мірою відповідний" застосовують рівнозначним чином в даному описі. Вони позначають фрагменти нуклеїнової кислоти, де зміни однієї або більше нуклеотидних основ не впливають на здатність фрагмента нуклеїнової кислоти опосередковувати експресію гена або продукувати певний фенотип. Ці терміни також позначають модифікації фрагментів нуклеїнових кислот за даним винаходом, такі як делеція або вставка одного або більше нуклеотидів, які істотною мірою не змінюють функціональних властивостей кінцевого фрагмента нуклеїнової кислоти в порівнянні з початковим, незміненим фрагментом. Таким чином слід розуміти, як буде зрозуміло фахівцю в даній галузі, що винахід охоплює більш ніж конкретні приклади послідовностей. Крім того, фахівцю в даній галузі буде зрозуміло, що істотною мірою подібні послідовності нуклеїнових кислот, включені до даного винаходу, також визначаються за їх здатністю гібридизуватися (в умовахпомірної жорсткості, наприклад, 0,5 x розчину цитрату і хлориду натрію [SSC], 0,1 % натрію додецилсульфату [SDS], 60 °C) з послідовностями, наведеними як приклади в даному описі, або з будь-якою частиною нуклеотидних послідовностей, розкритих в даному описі, і які є функціонально рівноцінними будь-якій з послідовностей нуклеїнових кислот, розкритих в даному описі. Суворість умов може бути скоректована для скринінгу помірною мірою подібних фрагментів, таких як гомологічні послідовності з віддаленою мірою споріднених організмів, до високою мірою подібних фрагментів, таких як гени, що подвоюють функціональні ферменти приблизно споріднених організмів. Промивання після гібридизації визначає жорсткість умов. Термін "селективно гібридизується" включає посилання на гібридизацію, в жорстких умовах гібридизації, послідовності нуклеїнової кислоти з вказаною цільовою послідовністю нуклеїнової кислоти до більшого ступеня, що піддається виявленню (наприклад, як мінімум вдвічі перевищує фон), в порівнянні з гібридизацією з нецільовими послідовностями нуклеїнових кислот, і до істотного виключення нецільових нуклеїнових кислот. Послідовності, що селективно гібридизуються, звичайно демонструють як мінімум приблизно 80 % ідентичність послідовності, або 90 % ідентичність послідовності, і, зокрема, до 100 % ідентичності (тобто, повністю комплементарні) послідовностей по відношенню одна до одної. Терміни "жорсткі умови" або "жорсткі умови гібридизації" означають посилання на умови, в яких зонд селективно гібридизується з цільовою послідовністю. Жорсткі умови залежать від послідовності і будуть різними за різних обставин. Шляхом контролю жорсткості гібридизації та/або умов промивання визначають цільові послідовності, що є на 100 % комплементарними по відношенню до зонда (гомологічне зондування). Альтернативно, жорсткість умов може бути скоректована таким чином, щоб дозволити деяку невідповідність послідовностей таким чином, що виявляються нижчі ступені подібності (гетерологічне зондування). Загалом, довжина зонду становить менш ніж приблизно 1000 нуклеотидів, необов'язково менш ніж 500 нуклеотидів. 13 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Звичайно, жорсткі умови будуть такими, при яких концентрація солі становить для іона Na менш ніж приблизно 1,5 M, типово концентрація іона Na (або інших солей) приблизно 0,01-1,0 M при pH 7,0-8,3, і температура становить як мінімум приблизно 30 °C для коротких зондів (наприклад, 10-50 нуклеотидів) і як мінімум приблизно 60 °C для довгих зондів (наприклад, більше 50 нуклеотидів). Суворі умови також можуть бути забезпечені додаванням дестабілізуючих агентів, таких як формамід. Приклади умов низької жорсткості включають гібридизацію з буферним розчином 30-35 % формаміду, 1 M NaCl, 1 % натрій додецилсульфату при 37 °C, і промивання в 1-2 x SSC (20 x SSC=3,0 M NaCl/0,3 M тринатрію цитрату) при 50-55 °C. Приклад умов помірної жорсткості включає гібридизацію при концентрації 40-45 % формаміду, 1 M NaCl, 1 % SDS при 37 °C, і промивання в 0,5-1 x SSC при 55-60 °C. Приклад умов високої жорсткості включає гібридизацію в 50 % формаміду, 1 M NaCl, 1 % SDS при 37 °C, і промивання в 0,1 x SSC при 60-65 °C. Специфічність звичайно є функцією промивання після гібридизації, причому критичними факторами розчину для кінцевого промивання є іонна сила і температура. Для гібридів ДНКДНК, Tm може бути наближено обчислена на базі рівняння Meinkoth et al., Anal. Biochem. 138:267-284 (1984): Tm=81.5 °C+16.6 (log M) + 0.41 (% GC) - 0.61 (% формаміду) - 500/L; де M - молярність одновалентних катіонів, % GC - відсоток нуклеотидів гуанозину і цитозину в ДНК, % формаміду - відсоток формаміду в розчині гібридизації, і L - довжина гібрида в парах основ. Tm - температура (при визначених значеннях іонної сили і pH), при якій 50 % цільової комплементарної послідовності гібридизуються з достеменно вирівняним зондом. T m знижується приблизно на 1 °C для кожного 1 % невідповідності; таким чином, T m, умови гібридизації та/або промивання можуть бути скоректовані для гібридизації з послідовностями бажаної ідентичності. Наприклад, якщо знайдені послідовності з ідентичністю 90 %, Tm може бути знижена на 10 °C. Загалом, жорсткі умови вибирають таким чином, щоб температура була приблизно на 5 °C нижча за температуру плавлення (Tm) для конкретної послідовності і комплементарної до неї при визначеній іонній силі і pH. Однак, при дуже жорстких умовах може використовуватися гібридизація та/або промивання при температурі на 1, 2, 3, або 4 °C нижче за температуру плавлення (Tm); при умовах помірної жорсткості може використовуватися гібридизація та/або промивання при температурі на 6, 7, 8, 9, або 10 °C нижче за температуру плавлення (Tm); при умовах низької жорсткості може використовуватися гібридизація та/або промивання при температурі на 11, 12, 13, 14, 15, або 20 °C, ніж температура плавлення (T m). Із застосуванням рівняння, гібридизації і складу рідини для промивання, а також бажаної T m, середній фахівець в даній галузі зрозуміє, що описані варіації жорсткості умов гібридизації та/або розчинів для промивання. Якщо бажаний ступінь невідповідності призводить до T m менше 45 °C (водний розчин) або 32 °C (розчин формаміду), переважним є підвищення концентрації SSC таким чином, щоб можна було застосовувати вищу температуру. Докладне керівництво з гібридизації нуклеїнових кислот міститься в Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier, New York (1993); і Current Protocols in Molecular Biology, Chapter 2, Ausubel et al., Eds., Greene Publishing and Wiley-Interscience, New York (1995). Умови гібридизації та/або промивання можуть бути застосовані протягом як мінімум 10, 30, 60, 90, 120 або 240 хвилин. "Ідентичність послідовностей" або "ідентичність" в контексті послідовностей нуклеїнових кислот або поліпептидів позначає основи нуклеїнової кислоти або залишки амінокислот в двох послідовностях, які є однаковими при вирівнюванні для максимальної відповідності у вказаному вікні порівняння. Таким чином, "відсоток ідентичності послідовності" позначає значення, визначене шляхом порівняння двох оптимально вирівняних послідовностей в межах вікна порівняння, де частина послідовності полінуклеотиду або поліпептиду у вікні порівняння може включати вставки або делеції (тобто, проміжки) в порівнянні з довідковою послідовністю (яка не містить вставок або делецій) для оптимального вирівнювання двох послідовностей. Відсоток обчислюють, визначаючи кількість положень, в яких розташовані ідентичні основи нуклеїнової кислоти або залишки амінокислот в обох послідовностях, з одержанням кількості відповідних положень, поділом кількості відповідних положень на загальну кількість положень у вікні порівняння та помноження результатів на 100 для одержання відсотка ідентичності послідовностей. Придатні приклади відсотка ідентичності послідовностей включають, не обмежуючись ними, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 % або 95 %, або будь-який цілочисельнний відсоток від 50 % до 100 %. Така ідентичність може бути визначена із застосуванням будь-якої з програм, описаних в даному описі. 14 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Вирівнювання послідовностей та обчислення відсотка ідентичності або подібності може бути здійснене із застосуванням різноманітних способів порівняння, розроблених для виявлення гомологічних послідовностей, в тому числі, не обмежуючись ними, програма MegAlign обчислювального набору для біоінформатики LASERGENE (DNASTAR Inc., Медісон, Вісконсин). В межах контексту даної заявки слід розуміти, що у випадку застосування з метою аналізу програмного забезпечення для аналізу послідовностей, результати аналізу будуть базуватися на "параметрах за умовчанням" з посиланням на програму, якщо не вказано інше. В даному описі "параметри за умовчанням" позначають будь-який набір значень або параметрів, які завантажуються при встановленні програмного забезпечення. "Спосіб вирівнювання Clustal V" відповідає способу вирівнювання під назвою Clustal V (описаний Higgins and Sharp, CABIOS. 5:151-153 (1989); Higgins, D.G. et al. (1992) Comput. Appl. Biosci. 8:189-191) і закладений в програму MegAlignобчислювального набору для біоінформатики LASERGENE (DNASTAR Inc., Медісон, Вісконсин). Для множинних вирівнювань, параметри за умовчанням відповідають наступним: штраф проміжку =10 і штраф довжини проміжку =10. Параметри, які використовують за умовчанням для попарного вирівнювання та обчислення відсотка ідентичності послідовностей білка із застосуванням способу Clustal, є наступними: KTUPLE = 1, штраф проміжку =3, вікно = 5 і збережені діагоналі = 5. Для нуклеїнових кислот параметри є наступними: KTUPLE = 2, штраф проміжку = 5, вікно = 4 і збережені діагоналі = 4. Після вирівнювання послідовностей із застосуванням програми Clustal V можна одержати "відсоток ідентичності" на базі таблиці "відстані послідовностей" в тій же програмі. "Спосіб вирівнювання BLASTN" представляє алгоритм, запропонований Національним центром біотехнологічної інформації (NCBI) для порівняння нуклеотидних послідовностей із застосуванням параметрів, які використовують за умовчанням. Фахівцю в даній галузі буде зрозуміло, що численні рівні ідентичності послідовностей є придатними для ідентифікації поліпептидів з інших видів, де такі поліпептиди володіють однаковою або подібною функцією або активністю. Придатні приклади відсотка ідентичності включають, не обмежуючись ними, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 % або 95 %, або будь-який цілочисельнний відсоток від 50 % до 100 %. Дійсно, будь-який цілочисельнний відсоток ідентичності амінокислот від 50 % до 100 % може бути придатним для опису даного винаходу, наприклад, 51 %, 52 %, 53 %, 54 %, 55 %, 56 %, 57 %, 58 %, 59 %, 60 %, 61 %, 62 %, 63 %, 64 %, 65 %, 66 %, 67 %, 68 %, 69 %, 70 %, 71 %, 72 %, 73 %, 74 %, 75 %, 76 %, 77 %, 78 %, 79 %, 80 %, 81 %, 82 %, 83 %, 84 %, 85 %, 86 %, 87 %, 88 %, 89 %, 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 % або 99 %. Також, представляє інтерес будь-яка послідовність, з повною довжиною або її частина, що є комплементарною до даного виділеного нуклеотидного фрагмента. "Ген" позначає фрагмент нуклеїнової кислоти, який експресує специфічний білок, зокрема, регуляторні послідовності, розташовані до (5’ некодуючі послідовності) і після (3’ некодуючі послідовності) кодуючої послідовності. "Природний ген" позначає ген, який знайдено в природі, з його власними регуляторними послідовностями. "Химерний ген" позначає будь-який ген, який не є природним геном, що включає регуляторні і кодуючі послідовності, які не знайдені разом в природі. Відповідно, химерний ген може включати регуляторні послідовності і кодуючі послідовності, які походять з різних джерел, або регуляторні послідовності і кодуючі послідовності, що походять з одного і того ж самого джерела, але організовані до деякої міри іншим чином, ніж знайдені в природі. "Чужорідний" ген позначає ген, який звичайно не знаходять в організмі-хазяїні, але який введений в організм-хазяїн переносом гена. Чужорідні гени можуть включати природні гени, вставлені в неприродний організм, або химерні гени. "Трансген" представляє ген, який введений в геном процедурою трансформації. Термін "геном" стосовно рослинної клітини включає не тільки хромосомну ДНК, знайдену в межах ядра, але і ДНК органел, знайдену в межах субклітинних компонентів (наприклад, мітохондрія, пластида) клітини. "Оптимізований за кодонами ген" представляє ген, сконструйований за частотою використання кодонів таким чином, щоб імітувати частоту переважного використання кодонів клітини-хазяїна. "Алель" - одна з декількох альтернативних форм гена, що займає даний локус на хромосомі. Якщо всі алелі, присутні в даному локусі на хромосомі, є однаковими, рослина є гомозиготною в зазначеному локусі. Якщо алелі, присутні в даному локусі на хромосомі, відрізняються, рослина є гетерозиготною в даному локусі. "Кодуюча послідовність" позначає послідовність ДНК, що кодує конкретну послідовність амінокислот. "Регуляторні послідовності" позначають нуклеотидні послідовності, розміщені в 15 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 напрямку 3’-5’ (5’ некодуючі послідовностей), в межах, або в напрямку 5’-3’ (3’ некодуючі послідовності) по відношенню до кодуючої послідовності, і які впливають на транскрипцію, обробку або стабільність РНК, або трансляцію супутньої кодуючої послідовності. Регуляторні послідовності можуть включати, не обмежуючись ними: промотори, лідерні послідовності трансляції, інтрони, послідовності розпізнавання поліаденілювання, сайти обробки РНК, сайти зв'язування з ефектором і стовбурово-петльові структури. "Промотор" позначає послідовність ДНК, здатну до управління експресією кодуючої послідовності або функціональної РНК. Послідовність промотору складається з проксимальних і більш дистальних в напрямку 3’-5’ елементів, останні елементи часто позначають як енхансери. Відповідно, "енхансер" є послідовністю ДНК, яка може стимулювати активність промотору, і може бути природним елементом промотору або гетерологічним елементом, вставленим таким чином, щоб збільшити рівень промотору або його специфічність по відношенню до тканин. Промотори можуть бути одержані у всій їх повноті з природного гена, або складатися з різних елементів, що походять з різних промоторів, знайдених в природі, або навіть включають синтетичні сегменти ДНК. Фахівцю в даній галузі буде зрозуміло, що різні промотори можуть спрямовувати експресію гена в різних типах тканин або клітин, або на різних стадіях розвитку, або у відповідь на різні природні умови. Крім того, визнано, що оскільки в більшості випадків точні межі регуляторних послідовностей повністю не визначені, деякі варіанти фрагментів ДНК можуть володіти ідентичною активністю промотору. Промотори, які примушують ген експресуватися в більшості типів клітин більшу частину часу, звичайно позначають як "конститутивні промотори". Постійно виявляють нові промотори різних типів, придатні для рослинних клітин; численні приклади можна знайти в компіляції Okamuro, J. K., and Goldberg, R. B. Biochemistry of Plants 15:1-82 (1989). "Лідерна послідовність трансляції" позначає полінуклеотидну послідовність, розміщену між послідовністю промотору гена і кодуючою послідовністю. У лідерній послідовності трансляції присутня повністю оброблена мРНК, розташована в напрямку 3’-5’ по відношенню до послідовності ініціації трансляції. Лідерна послідовність трансляції може впливати на обробку первинного транскрипта до мРНК, стабільність мРНК або ефективність трансляції. Описані приклади лідерних послідовностей трансляції (Turner, R. and Foster, G. D., Mol. Biotechnol. 3:225-236 (1995)). "3’ некодуючі послідовності", "термінатор транскрипції" або "термінаторні послідовності" позначають послідовності ДНК, що розміщені в напрямку 5’-3’ по відношенню до кодуючої послідовності, і включають послідовності розпізнавання поліаденілювання та інші послідовності, що кодують регуляторні сигнали, здатні впливати на обробку мРНК або експресію гена. Сигнал поліаденілювання звичайно характеризується впливом на додавання ділянок поліаденілової кислоти до 3’ кінця прекурсора мРНК. Приклади застосування різних 3’ некодуючих послідовностей наведені в Ingelbrecht, I. L., et al. Plant Cell 1:671-680 (1989). "Транскрипт РНК" позначає продукт, одержаний в результаті каталізованої РНКполімеразою транскрипції послідовності ДНК, що. Якщо транскрипт РНК представляє довершену додаткову копію послідовності ДНК, його позначають як первинний транскрипт. Транскрипт РНК позначають як зрілу РНК, коли він являє собою послідовність РНК, що є результатом посттранскрипційної обробки первинного транскрипта. "Інформаційна (матрична) RNA" або "мРНК" позначає РНК який без інтронів, яка може бути трансльована клітиною в білок. "кДНК" позначає ДНК, яка є комплементарною до і синтезована з шаблону мРНК із застосуванням ферменту зворотною транскриптази. кДНК може бути одноланцюговою або може бути перетворена на двохланцюгову із застосуванням фрагмента Кленоу (Klenow) ДНКполімерази І. "Смислова" РНК позначає транскрипт РНК, який включає мРНК і може бути трансльований в білок в межах клітини або in vitro. "Антисмислова РНК" позначає транскрипт РНК, який є комплементарний до всієї або частини цільового первинного транскрипта або мРНК, і яка блокує експресію цільового гена (патент США 5,107,065). Комплементарність антисмислової РНК може стосуватися будь-якої частини конкретного транскрипта гена, тобто, 5’ некодуючої послідовності, 3’ некодуючої послідовності, інтронів або кодуючої послідовності. "Функціональна РНК" позначає антисмислову РНК, рибозимну РНК або іншу РНК, яка не може бути трансльована, але все ще здійснює вплив на клітинні процеси. Терміни "комплементарний" і "зворотно комплементарний" застосовують рівнозначним чином в даному описі щодо транскриптів мРНК, і вони призначені визначити антисмислову РНК одиниці генетичного коду. Термін "функціонально зв'язаний" позначає сполучення послідовностей нуклеїнових кислот на єдиному фрагменті нуклеїнової кислоти таким чином, що функція одного регулюється іншим. Наприклад, промотор є функціонально зв'язаним з кодуючої послідовністю, якщо він здатний регулювати експресію зазначеної кодуючої послідовності (тобто, кодуюча послідовність 16 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 знаходиться під транскрипційним контролем промотору). Кодуючі послідовності можуть бути функціонально зв'язані з регуляторними послідовностями в смисловій або антисмисловій орієнтації. В іншому прикладі, комплементарні ділянки РНК за винаходом можуть бути функціонально зв'язані, безпосередньо або непрямо, в положенні 5’ з цільовою мРНК, або в положенні 3’ з цільовою мРНК, або в межах цільової мРНК, або перша комплементарна ділянка розташована в положенні 5’, і комплементарна до неї ділянка - в положенні 3’ цільової мРНК. Стандартна рекомбінантна ДНК і молекулярні методи клонування, використані в даному описі, добре відома в даній галузі і описані більш детально в Sambrook, J., Fritsch, E.F. and Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY (1989). Способи трансформації добре відомі фахівцям в даній галузі і описані нижче. "ПЛР" або "полімеразна ланцюгова реакція" є технікою синтезу великих кількостей конкретних сегментів ДНК і складається з серії циклів, що повторюються (Perkin Elmer Cetus Instruments, Norwalk, CT). Звичайно, двохланцюгову ДНК денатурують нагріванням, два праймери, комплементарні до 3’ меж цільового сегменту, відпалюють при низькій температурі, після чого подовжують при проміжній температурі. Один набір вказаних трьох послідовних стадій позначається як "цикл". Термін "рекомбінантний" позначає штучну комбінацію двох іншим чином розділених сегментів послідовності, наприклад, хімічним синтезом або маніпулюванням виділеними сегментами нуклеїнових кислот за допомогою методів генної інженерії. Терміни "плазміда", "вектор" і "касета" позначають позахромосомний елемент, який часто несе гени, що не є частиною центрального метаболізму клітини, звичайно у формі дугових двохланцюгових фрагментів ДНК. Такі елементи можуть бути послідовностями, що реплікуються автономно, послідовностями, що інтегруються в геном, послідовностями фага або нуклеотидними послідовностями, лінійними або дуговими, одно- або двохланцюговими ДНК або РНК, одержаними з будь-якого джерела, в якому цілий ряд нуклеотидних послідовностей з'єднаний або рекомбінований в унікальний конструкт, здатний до введення промоторного фрагмента і послідовності ДНК для вибраного генного продукту, разом з відповідною 3' нетрансльованою послідовністю, в клітину. "Трансформаційна касета" позначає конкретний вектор, що містить чужорідний ген і на додаток до чужорідного гена містить елементи, які полегшують трансформацію конкретної клітини-хазяїна. "Касета експресії" позначає конкретний вектор, що містить чужорідний ген і на додаток до чужорідного гена містить елементи, які дозволяють збільшену експресію такого гена в чужорідному хазяїні (тобто, до дискретного фрагмента містить елементи, в який може бути переміщена послідовність або фрагмент нуклеїнової кислоти). Терміни "рекомбінантний конструкт", "конструкт експресії", "химерний конструкт", "конструкт" і "рекомбінантний конструкт" ДНК застосовують в даному описі рівнозначним чином. Рекомбінантний конструкт включає штучну комбінацію фрагментів нуклеїнових кислот, наприклад, регуляторних і кодуючих послідовностей, які не знайдені разом в природі. Наприклад, химерний конструкт може включати регуляторні послідовності і кодуючі послідовності, які походять з різних джерел, або регуляторні послідовності і кодуючі послідовності, що походять з одного джерела, але організовані до деякої міри іншим чином, ніж знайдено в природі. Такий конструкт може бути використаний окремо або може бути використаний в з'єднанні з вектором. Якщо застосовується вектор, то вибір вектора залежить від способу, який буде застосовуватися для перетворення клітин-хазяїв, як добре відомо фахівцям в даній галузі. Наприклад, може бути використаний плазмідний вектор. Кваліфікований спеціаліст добре поінформований про генетичні елементи, які повинні бути присутні на векторі для успішного перетворення, селекції та розведення клітин-хазяїв, що включають будь-який з виділених фрагментів нуклеїнових кислот за винаходом. Кваліфікований фахівець також зрозуміє, що різні незалежні явища трансформації будуть приводити до різних рівнів і характеру експресії (Jones et al., EMBO J. 4:2411-2418 (1985); De Almeida et al., Mol. Gen. Genetics 218:78-86 (1989)), і таким чином, що необхідно здійснити скринінг множинних подій для одержання ліній, що демонструють бажаний рівень і характер експресії. Такий скринінг може бути здійснений, серед інших, Саузерн-аналізом ДНК, Нозерн-аналізом експресії мРНК, аналізом експресії білка методом імуноблотингу або фенотипічним аналізом. Термін "експресія" в даному описі позначає продукування функціонального кінцевого продукту (наприклад, мРНК або білок [прекурсор або зрілий]). Термін "введений" позначає введення нуклеїнової кислоти (наприклад, конструкт експресії) або білка в клітину. "Введений" включає посилання на інкорпорацію нуклеїнової кислоти в еукаріотну або прокаріотну клітину, де нуклеїнова кислота може бути інкорпорована в геном клітини, і включає посилання на тимчасове введення нуклеїнової кислоти або білка в клітину. 17 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 "Введений" включає посилання на способи стабільної або тимчасової трансформації, а також статеве схрещення. Таким чином, "введений" в контексті вставки фрагмента способи (наприклад, рекомбінантний конструкт ДНК/конструкт експресії) в клітину позначає "трансфекцію" або "трансформацію" або "перетворення" і включає посилання на інкорпорацію фрагмента нуклеїнової кислоти в еукаріотну або прокаріотну клітину, де фрагмент нуклеїнової кислоти може бути інкорпорований в геном клітини (наприклад, хромосомна, плазмідна, пластидна або мітохондріальна ДНК), перетворений на автономний реплікон або тимчасово експресований (наприклад, трансфікована мРНК). "Зрілий" білок позначає посттрансляційно оброблений поліпептид (тобто, такий, з якого видалені будь-які пре- або пропептиди, присутні в первинному продукті трансляції). Білок"прекурсор" позначає первинний продукт трансляції мРНК (тобто, в якому все ще присутні пре- і пропептиди). Пре- і пропептиди можуть являти собою, не обмежуючись ними, сигнали внутрішньоклітинної локалізації. "Стабільна трансформація" позначає перенос фрагмента нуклеїнової кислоти в геном клітини-хазяїна, в тому числі як ядерний, так і органеллярний геноми, що дає генетично стабільну спадковість. На протилежність цьому, "тимчасова трансформація" позначає перенос фрагмента нуклеїнової кислоти в ядро або органелу, що містить ДНК, організму-хазяїна, що приводить до експресії гена без інтеграції або стабільної спадковості. Організми-хазяї, що містять трансформовані фрагменти нуклеїнової кислоти, позначають як "трансгенні" організми. В даному описі "трансгенний" позначає рослину або клітину, яка містить в геномі гетерологічний полінуклеотид. Переважно, гетерологічний полінуклеотид стабільно інкорпорований в геном таким чином, що полінуклеотид передається наступним поколінням. гетерологічний полінуклеотид може бути інкорпорований в геном окремо або як частина конструкта експресії. "Трансгенний" в даному описі включає будь-яку клітину, лінію клітин, калюс, тканину, частину рослини або рослину, генотип якої змінений присутністю гетерологічної нуклеїнової кислоти, в тому числі трансгенні особини, початково модифіковані таким чином, а також одержані статевим схрещенням або безстатевим розмноженням з початкової трансгенної особини. Термін "трансгенний" в даному описі не включає модифікацію генома (хромосомного або позахромосомного) звичайними методами розведення рослин або природними явищами, такими як випадкове перехресне запліднення, інфікування нерекомбінантними вірусами, трансформація нерекомбіннатними бактеріями, нерекомбінантна транспозиція або спонтанна мутація. "Антисмислове інгібування" позначає продукування транскриптів антисмислової РНК, здатних до пригнічення експресії цільового білка. "Співпригнічення" позначає продукування смислових транскриптів РНК, здатних до пригнічення експресії ідентичних або істотною мірою подібних чужорідних або ендогенних генів (патент США 5,231,020). Конструкти співпригнічення в рослинах раніше створювали, зосереджуючись на надмірній експресії послідовності нуклеїнової кислоти, що є гомологічною до ендогенної мРНК, в смисловій орієнтації, що приводило до зниження рівнів всіх РНК, гомологічних до надмірно експресованої послідовності (Vaucheret et al., Plant J. 16:651-659 (1998); Gura, Nature 404:804-808 (2000)). Загальна ефективність даного феномена є низькою, і ступінь зниження рівнів РНК варіює в широких межах. В нещодавній роботі описане застосування структур "шпильки", які інкорпорують всю або частину кодуючої послідовності мРНК, в комплементарній орієнтації, що приводить до потенційної "стовбуровопетльової" структури для експресованої РНК (PCT публікація WO 99/53050, опублікована 21 жовтня 1999 р.; PCT публікація WO 02/00904 опублікована 3 січня 2002 р.). Це збільшує частоту співпригнічення у регенеарованих трансгенних рослинах. Інша варіація описує застосування послідовностей рослинних вірусів для спрямування пригнічення або "сайленсингу", проксимальних кодуючих послідовностей мРНК (PCT публікація WO 98/36083, опублікована 20 серпня 1998 р.). Обидва описані феномени співпригнічення прояснені механістично, хоча генетичні факти почала проливати світло на цю складну ситуацію (Elmayan et al., Plant Cell 10:1747-1757 (1998)). Термін "олійний" позначає такі організми, які прагнуть зберігати джерело енергії у формі ліпідів (Weete, у: Fungal Lipid Biochemistry, 2nd Ed., Plenum, 1980). Клас рослин, ідентифікованих як олійні, звичайно позначають як рослини "олійної культури". Приклади рослин олійної культури включають, не обмежуючись ними: сою (види Glycine і Soja), льон (види Linum), рапс (види Brassica), маїс, бавовну, сафлор (види Carthamus) і соняшник (види Helianthus). В олійних мікроорганізмах клітинна олія або вміст ТАГ загалом відповідає сигмоподібній кривій, де концентрація ліпіду зростає до максимуму в пізній логарифмічній або ранній стаціонарній фазі росту, після чого поступово зменшується протягом пізньої стаціонарної фази і фази загибелі (Yonгanitchai and Ward, Appl. Environ. Microbiol. 57:419-25 (1991)). Термін 18 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 "олеогені дріжджі" позначає такі мікроорганізми, класифіковані як дріжджі, які виробляють олію. Для олеогених мікроорганізмів досить звичайним є надлишкове накопичення приблизно 25 % сухої маси клітини у вигляді олії. Приклади олеогених дріжджів включають, не обмежуючись ними, наступні видів: Yarrowia, Candida, Rhodotorula, Rhodosporidium, Cryptococcus, Trichosporon і Lipomyces. Термін "Euglenophyceae" позначає групу одноклітинних, безбарвних або здатних до фотосинтезу джгутикових ("евгленові"), знайдених в прісноводних та морських водоймищах, землі і паразитичних середовищах. Клас характеризується одинарними одноклітинними організмами, більшість з яких вільно плавають і мають два джгутики (один з яких може бути не емергентним), що ростуть з передньої інвагінації, відомої як резервуар. Здатні до фотосинтезу евгленові містять від одного до декількох хлоропластів кольору зеленої трави, які варіюють від незначних дисків до поширених пластин або стрічок. Безбарвні евгленові залежать від осмотрофії або фаготрофії для асиміляції поживних речовин. Близько 1000 видів описано і класифіковано приблизно в 40 родах і 6 підкласах. Приклади Euglenophyceae включають, не обмежуючись ними, наступні види: Eutreptiella і Tetruetreptia. Термін "рослина" позначає суцільні рослини, органи рослини, тканини рослин, зерна, клітини рослин, а також їх зерна і нащадка. Клітини рослин включають, не обмежуючись ними, клітини зерен, суспензійні культури, ембріони, меристематичні ділянки, тканини калюсу, листя, коріння, бруньки, гаметофіти, спорофіти, пилок і мікроспори. "Нащадок" включає будь-яке подальше покоління рослини. Короткий огляд: мікробний біосинтез жирних кислот і триацилгліцеридів Загалом, акумуляція ліпідів в олеогених мікроорганізмах, що розпочинається у відповідь на загальне співвідношення вуглецю до азоту в середовищі росту. Даний процес, що приводить до синтезу de novo вільного пальмітату (16:0) в олеогених мікроорганізмах, детально описаний в PCT публікації WO 2004/101757. Пальмітат є прекурсором похідних насичених і ненасичених жирних кислот з довшим ланцюгом, які утворюються під дією елонгаз і десатураз (фіг. 6). ТАШ (первинна одиниця зберігання для жирних кислот) утворюються в результаті серії реакцій, які включають: (1) естерифікацію однієї молекули ацил-КоА до гліцерин-3-фосфату під дією ацилтрансферази з утворенням лізофосфатидинової кислоти; (2) естерифікацію другої молекули ацил-КоА під дією ацилтрансферази з утворенням 1,2-діацилгліцерид фосфату (звичайно ідентифікований як фосфатидна кислота); (3) видалення фосфату фосфатазою фосфатидної кислоти з утворенням 1,2-діацилгліцериду (DAG); і (4) приєднання третьої жирної кислоти під дією ацилтрансферази з утворенням ТАГ. Широкий спектр жирних кислот може бути введений в ТАГ, зокрема, насичені і ненасичені жирні кислоти та жирні кислоти з коротким і довгим ланцюгом. Біосинтез омега-жирних кислот Метаболічний процес, в якому олеїнова кислота перетворюється на омега-3/омега-6 жирні кислоти з довгим ланцюгом включає подовження вуглецевого ланцюга додаванням вуглецевих атомів і забезпечення ненасиченості молекули через утворення подвійних зв'язків. Це вимагає серії спеціальних ферментів для та утворення ненасиченості в мембрані ендоплазматичного ретикулума. Однак, як можна побачити на фіг. 6 і як описано нижче, часто існують множинні альтернативні шляхи продукування конкретної омега-3/омега-6 жирної кислоти з довгим ланцюгом. Конкретно, всі шляхи вимагають початкового перетворення олеїнової кислоти на ЛК, першої з омега-6 жирних кислот, під дією дельта-12 десатурази. Далі, із застосуванням "шляху дельта9 елонгази/дельта-8 десатурази", омега-6 жирні кислоти з довгим ланцюгом утворюються, як показано нижче: (1) ЛК перетворюється на ЕДК під дією дельта-9 елонгази; (2) ЕДК перетворюється на ДГЛК під дією дельта-8 десатурази; і (3) ДГЛК перетворюється на АРК під дією дельта-5 десатурази. Альтернативно, "шлях дельта-9 елонгази/дельта-8 десатурази" може використовуватися для утворення омега-3 жирних кислот з довгим ланцюгом, як вказано нижче: (1) ЛК перетворюється на AЛК, першу з омега-3 жирних кислот, під дією дельта-15 десатурази; (2) AЛК перетворюється на ЕТрК під дією дельта-9 елонгази; (3) ЕТрК перетворюється на ЕТК під дією дельта-8 десатурази; (4) ЕТК перетворюється на ЕПК під дією дельта-5 десатурази; (5) ЕПК перетворюється на ДПК під дією C20/22 елонгази; і (6) ДПК перетворюється на ДГК під дією дельта-4 десатурази. Необов'язково, омега-6 жирні кислоти можуть бути перетворені на омега-3 жирні кислоти; наприклад, ЕТК і ЕПК утворюються з ДГЛК і АРК, відповідно, під дією дельта-17 десатурази. В альтернативних шляхах біосинтезу омега-3/омега-6 жирних кислот використовується дельта-6 десатураза і C18/20 елонгаза (також відома як дельта-6 елонгаза, терміни можуть застосовуватися рівнозначним чином) (тобто, "шлях дельта-6 десатурази/дельта-6 елонгази"). 19 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 Більш конкретно, ЛК і AЛК можуть бути перетворені на ГЛК і СТК, відповідно, під дією дельта-6 десатурази; далі, C18/20 елонгази перетворює ГЛК на ДГЛК та/або СТК до ЕТК. Передбачається, що конкретні функціональні одиниці, необхідні для введення в конкретний організм-хазяїн для продукування омега-3/омега-6 жирних кислот, будуть залежати від клітинихазяїна (і його природного профілю ПНЖК та/або профілю десатурази/елонгази), доступності субстрату і бажаного кінцевого продукту(ів). Наприклад, експресія шляху дельта-9 елонгази/дельта-8 десатурази може бути переважною в деяких варіантах, на протилежність експресії шляху дельта-6 десатурази/дельта-6 елонгази, оскільки ПНЖК, продуковані через перший шлях, не містять ГЛК. Фахівець в даній галузі зможе ідентифікувати різні гени-кандидати, що кодують кожний з ферментів, бажаних для омега-3/омега-6 біосинтезу жирних кислот. Придатні послідовності десатурази та елонгази можуть походити з будь-якого джерела, наприклад, виділений з природного джерела (бактерій, морських водоростей, грибів, рослин, тварин і т. п.), продукований напівсинтетичним способом або синтезований de novo. Хоча конкретне джерело генів десатурази і елонгази, введених в хазяїна, не критичне, фактори для вибору конкретного поліпептиду, що володіє активністю десатурази або елонгази, включають: (1) специфіку субстрату поліпептиду; (2) чи є поліпептид або його компонент обмежуючим швидкість ферментом; (3) чи є десатураза або елонгаза незамінною для синтезу бажаної ПНЖК; та/або (4) кофактори, яких вимагає поліпептид. Поліпептид, що експресується, переважно має параметри, сумісні з біохімічним середовищем, що оточує його в клітині-хазяїні (див. PCT публікацію WO 2004/101757 щодо подробиць). У додаткових варіантах, також буде корисним розглянути ефективність перетворення кожної конкретної десатурази та/або елонгази. Більш конкретно, оскільки кожен фермент рідко функціонує із 100 % ефективністю для перетворення субстрату на продукт, кінцевий ліпідний профіль неочищеної олії, продукованой в клітині-хазяїні, звичайно буде демонструвати суміш різних ПНЖК, що складається з бажаної омега-3/омега-6 жирної кислоти, а також різноманітних проміжних ПНЖК, що передують їй в шляху біосинтезу. Таким чином, розгляд ефективності перетворення кожного ферменту також представляє змінну для оптимізації біосинтезу бажаної жирної кислоти, яка повинна розглядатися в світлі бажаного кінцевого ліпідного профілю продукту. Враховуючи кожен з наведених вище факторів, гени-кандидати, що володіють відповідною активністю десатурази та елонгази (наприклад, дельта-6 десатурази, C18/20 елонгази, дельта-5 десатурази, дельта-17 десатурази, дельта-15 десатурази, дельта-9 десатурази, дельта-12 десатурази, C14/16 елонгази, C16/18 елонгази, дельта-9 елонгази, дельта-8 десатурази, дельта-4 десатураза і C20/22 елонгази), можуть бути ідентифіковані у відповідності до публічно доступної літератури (наприклад, GenBank), патентної літератури та експериментального аналізу організмів, здатних до продукування ПНЖК. Такі гени будуть придатними для введення в конкретний організм-хазяїн, щоб дозволити або збільшити синтез ПНЖК організмом. Ідентифікація послідовності нової дельта-8 десатурази У даному винаході, нуклеотидні послідовності, що кодують дельта-8 десатурази, виділені з Tetruetreptia pomquetensis CCMP1491 (позначена в даному описі як "TpomD8"), виду Eutreptiella CCMP389 (позначений в даному описі як "E389D8") і Eutreptiella cf_gymnastica CCMP1594 (позначений в даному описі як "E1594D8"). Таким чином, даний винахід стосується виділеного полінуклеотиду, який включає: (a) нуклеотидну послідовність, що кодує поліпептид, який володіє активністю дельта-8 десатурази, де поліпептид демонструє як мінімум 80 % ідентичність амінокислот, за даними вирівнювання способом Clustal V, в порівнянні з послідовністю амінокислот, представленою в SEQ ID NO: 47 [E389D8], SEQ ID NO: 49 [E1594D8] або SEQ ID NO: 57 [TpomD8]; (b) нуклеотидну послідовність, що кодує поліпептид, який володіє активністю дельта-8 десатурази, де нуклеотидна послідовність демонструє як мінімум 80 % ідентичність послідовності, за даними вирівнювання способом BLASTN, в порівнянні з нуклеотидною послідовністю, представленою в SEQ ID NO: 92 [E389D8], SEQ ID NO: 93 [E1594D8] або SEQ ID NO: 62 [TpomD8]; або, (c) послідовність, комплементарну послідовності (a) нуклеотиду або (b), де комплементарна і нуклеотидна послідовності складаються з однакової кількості нуклеотидів і є на 100 % комплементарними. В іншому аспекті, даний винахід стосується виділеного полінуклеотиду, що включає нуклеотидну послідовність, яка кодує поліпептид, що володіє активністю дельта-8 десатурази, де нуклеотидна послідовність демонструє як мінімум 90 % ідентичність послідовності, за 20 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 даними вирівнювання способом BLASTN, в порівнянні з нуклеотидною послідовністю, представленою в SEQ ID NO: 92, SEQ ID NO: 93 або SEQ ID NO: 62. У альтернативних варіантах, необхідні послідовності десатурази E389D8, E1594D8 або TpomD8 можуть бути оптимізовані за кодонами для експресії в конкретному організмі-хазяїні. Як добре відомо в даній галузі, це може бути придатним засобом для додаткової оптимізації експресії ферменту в альтернативному хазяїні, оскільки застосування переважних для хазяїна кодонів може істотною мірою збільшити експресію чужорідного гена, що кодує поліпептид. Загалом, переважні для хазяїна кодони можуть бути визначені в межах конкретних цільових видів хазяїв, шляхом розгляду використання кодону в білках (переважно таких, що експресуються у найбільших кількостях) і визначенням кодонів, які використовуються з найбільшою частотою. Далі, кодуюча послідовність для цільового поліпептиду, що володіє, наприклад, активністю десатурази, може бути синтезована в цілому або частково, із застосуванням кодонів, переважних у видах хазяїв. В одному варіанті винаходу, описаного в даному описі, E389D8, E1594D8 та/або TpomD8 можуть бути оптимізовані за кодонами для експресії в Yarrowia lipolytica, як викладено в PCT публікації WO 04/101757. В альтернативних варіантах, може бути бажаним змінити частину кодонів, що кодують E389D8, E1594D8 та/або TpomD8 (як показано SEQ ID NO: 92, 93 і 62, відповідно) для збільшення експресії гена в організмі-хазяїні, в тому числі, не обмежуючись ними, рослині або частині рослини. Фахівець в даній галузі може використати розкриття даного опису для створення різних інших оптимізованих за кодонами білків дельта-8 десатурази, придатних для оптимальної експресії в альтернативних хазяях, на базі послідовностей дикого типу E389D8, E1594D8 та/або TpomD8. Відповідно, даний винахід стосується будь-якого оптимізованого за кодонами білка дельта-8 десатурази, який походить від E389D8 дикого типу (тобто, кодований SEQ ID NO: 47), E1594D8 дикого типу (тобто, кодований SEQ ID NO: 49) або TpomD8 дикого типу (тобто, кодований SEQ ID NO: 57). Ідентифікація та виділення гомологів Будь-яка з необхідних послідовностей десатураз (тобто, E389D8, E1594D8 або TpomD8) або їх частини можуть бути використані для пошуку гомологів дельта-8 десатурази в тому ж самому або іншому виді бактерій, водоростей, грибів, евгленові або рослини із застосуванням програмного забезпечення для аналізу послідовностей. Загалом, таке комп'ютерне програмне забезпечення підбирає подібні послідовності, відносячи ступінь гомології до різноманітних замін, делецій та інших модифікацій. Альтернативно, будь-яка з необхідних послідовностей десатураз або її частина також може бути використана як реагенти для гібридизації з метою ідентифікації гомолога дельта-8 десатурази. Основні компоненти тесту гібридизації нуклеїнової кислоти включають зонд, зразок з підозрюваним вмістом цільового гена або фрагмента гена і конкретний спосіб гібридизації. Зонди за даним винаходом звичайно являють собою послідовності одноланцюгових нуклеїнових кислот, комплементарні по відношенню до послідовностей нуклеїнових кислот, які підлягають виявленню. Зонди "здатні до гібридизації" з послідовностями нуклеїнових кислот, які підлягають виявленню. Хоча довжина зонда може варіювати від 5 основ до десятків тисяч основ, звичайно придатна довжина зонда становить від приблизно 15 основ до приблизно 30 основ. Тільки частина молекули зонда повинна бути комплементарною до послідовності нуклеїнової кислоти, яка підлягає виявленню. Крім того, немає необхідності в абсолютній компліментарності між зондом і цільовою послідовністю. Гібридизація відбувається між молекулами, що не є абсолютно комплементарними; в результаті частина основ в гібридизованій ділянці залишаються неспареними з відповідною комплементарною основою. Способи гібридизації добре відомі. Звичайно зонд і зразок необхідно змішати в умовах, які дозволяють гібридизацію нуклеїнових кислот. Це включає контакт зонда і зразка в присутності неорганічної або органічної солі при відповідних умов концентрації і температури. Зонд і зразок нуклеїнові кислоти повинні знаходитися в контакті достатньо довго, щоб дозволити будь-яку можливу гібридизацію між зондом і зразком нуклеїнових кислот. Концентрація зонда або мішені в суміші буде визначати час, необхідний для гібридизації. Чим вища концентрація зонда або мішені, тим менший час інкубації необхідний для гібридизації. Необов'язково, може бути доданий хаотропний агент (наприклад, гуанідінію хлорид, гуанідінію тіоціанат, натрію тіоціанат, літію тетрахлорацетат, натрію перхлорат, рубідію тетрахлорацетат, калію йодид, цезію трифторацетат). За бажанням, до суміші гібридизації може бути доданий формамід, звичайно 30-50 % (об./об.). Можуть використовуватися різні розчини гібридизації. Звичайно, вони містять від приблизно 20 до 60 % (об.), переважно 30 %, полярного органічного розчинника. Звичайний розчин для 21 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 гібридизації містить приблизно 30-50 % (об./об.) формаміду, приблизно 0,15-1 M хлориду натрію, приблизно 0,05-0,1 M буфера (наприклад, натрію цитрат, Трис-HCl, PIPES або HEPES [інтервал pH приблизно 6-9]), приблизно 0,05-0,2 % детергенту (наприклад, натрію додецилсульфату), або 0,5-20 мМ ЕДТА, FICOLL (Pharmacia Inc.) (приблизно 300-500 кДа), полівінілпіролідон (приблизно 250-500 кДа) і альбумін сироватки. Також типовий розчин для гібридизації містить немічені нуклеїнові кислоти носія (приблизно 0,1-5 мг/мл), фрагментована ядерна ДНК (наприклад, ДНК тимусу теляти або сперми лосося, або РНК дріжджів), і необов'язково приблизно 0,5-2 % (мас./об.) гліцину. Також можуть бути включені інші добавки, такі як агенти виключення об'єму, що включають різноманітні полярні водорозчинні засоби або засоби, що набрякають (наприклад, поліетиленгліколь), аніонні полімери (наприклад, поліакрилат або поліметакрилат) і аніонних полімери цукрів (наприклад, декстрану сульфат). Гібридизація нуклеїнових кислот може бути адаптована до різноманітних форматів аналізу. Одним з найбільш придатних є формат сендвічевого аналізу. Сендвічевий аналіз особливо може бути адаптований до гібридизації в не денатуруючих умовах. Основним компонентом аналізу сендвічевого типу є тверда основа. На твердій основі адсорбований або приєднаний до неї ковалентними зв'язками іммобілізований зонд нуклеїнової кислоти, немічений і комплементарний до однієї частини послідовності. В додаткових варіантах, будь-який з фрагментів нуклеїнових кислот дельта-8 десатурази, описаних в даному описі (або будь-який його ідентифікований гомолог), може бути використаним для виділення генів, що кодують гомологічні білки, з такого ж або іншого виду бактерій, водоростей, грибків, евгленових або рослин. Виділення гомологічних генів із застосуванням залежних від послідовності протоколів добре відома в даній галузі. Приклади залежних від послідовності протоколів включають, не обмежуючись ними: (1) способи гібридизації нуклеїнових кислот; (2) способи ампліфікації ДНК та РНК, наприклад, різноманітні способи використання технологій ампліфікації нуклеїнових кислот [наприклад, полімеразна ланцюгова реакція (ПЛР), Mullis et al., патент США 4,683,202; лігазна ланцюгова реакція (LCR), Tabor et al., Proc. Acad. Sci. USA 82:1074 (1985); або ампліфікація із зсувом ланцюга (SDA), Walker et al., Proc. Natl. Acad. Sci. U.S.A., 89:392 (1992)]; і (3) способи конструювання бібліотек та скринінг методом комплементації. Наприклад, гени, що кодують білки або поліпептиди, подібні до дельта-8 десатурази, описаної в даному описі, можуть бути виділені безпосередньо із застосуванням всіх або частини необхідних фрагментів нуклеїнових кислот як зондів гібридизації ДНК для скринінгу бібліотек, наприклад, з будь-яких цільових дріжджів або грибів, із застосуванням методології, добре відомої фахівцям в даній галузі (в якій організми, що продукують ДГЛК та/або ЕТК, будуть переважними). Конкретні олігонуклеотидні зонди на базі необхідних послідовностей нуклеїнових кислот можуть бути розроблені і синтезовані способами, відомими в даній галузі (Maniatis, вище). Крім того, повні послідовності можуть бути використані безпосередньо для синтезу зондів ДНК способами, відомими кваліфікованому фахівцю (наприклад, мічення ДНК випадковими праймерами, трансляція одноланцюгового розриву або методи кінцевої мітки), або зондів РНК, із застосуванням доступних систем транскрипції in vitro. Крім того, конкретні праймери можуть бути розроблені і застосовані для ампліфікації частини (або повної послідовності) необхідних послідовностей. В одержані продукти ампліфікації можуть бути введені мітки безпосередньо в ході реакцій ампліфікації або після реакцій ампліфікації, і вони можуть бути використані як зонди для виділення фрагментів ДНК повної довжини в умовах відповідної жорсткості. Звичайно, в методах ампліфікації по типу ПЛР, праймери мають різні послідовності і не є комплементарними один до одного. В залежності від бажаних умов, послідовності праймерів необхідно конструювати таким чином, щоб передбачити як ефективний, так і правильну реплікацію цільової нуклеїнової кислоти. ПЛР методи конструювання праймерів розповсюджені і добре відомі в даній галузі (Thein and Wallace, "The use of oligonucleotide as specific hybridization probes in the Diagnosis of Genetic Disorders", в Human Genetic Diseases: A Practical Approach, K. E. Davis Ed., (1986) pp 33-50, IRL: Herndon, VA; and Rychlik, W., в Methods in Molecular Biology, White, B. A. Ed., (1993) Vol. 15, pp 31-39, PCR Protocols: Current Methods and Applications. Humania: Totowa, NJ). Загалом два короткі сегменти необхідних послідовностей можуть бути використані в ПЛР протоколах для ампліфікації довших фрагментів нуклеїнової кислоти, що кодують гомологічні гени з ДНК або РНК. ПЛР також може бути здійснена на бібліотеці клонованих фрагментів нуклеїнових кислот, де послідовність одного праймера походить від необхідних фрагментів нуклеїнових кислот, і в послідовності іншого праймера використовується перевага присутності ділянок поліаденілової кислоти до 3' кінця прекурсора мРНК, що кодує еукаріотні гени. 22 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Альтернативно, друга послідовність праймера може базуватися на послідовностях, що походять від вектора клонування. Наприклад, кваліфікований фахівець може дотримуватися протоколу RACE (Frohman et al., PNAS USA 85:8998 (1988)) для генерації кДНК із застосуванням ПЛР, щоб ампліфікувати копії ділянки між єдиною точкою в транскрипті і 3' або 5' кінцем. Праймери, орієнтовані в 3' і 5' напрямках, можуть бути сконструйовані на базі необхідних послідовностей. З використанням комерційно доступних систем 3' RACE або 5' RACE (Gibco/BRL, Gaithersburg, MD) можуть бути виділені специфічні 3' або 5' фрагменти кДНК (Ohara et al., PNAS USA 86:5673 (1989); Loh et al., Science 243:217 (1989)). В інших варіантах, будь-який з фрагментів нуклеїнових кислот дельта-8 десатурази, описаних в даному описі (або будь-який їх ідентифікований гомолог) може бути використаний для створення нових, покращених десатураз жирних кислот. Як добре відомо в даній галузі, мутагенезі і селекція in vitro, хімічний мутагенез, способи "перестановки генів" або інші засоби можуть бути використані для введення мутацій в природні гени десатураз. Альтернативно, покращені жирні кислоти можуть бути синтезовані перестановкою доменів, де функціональний домен з будь-якого фрагмента нуклеїнових кислот дельта-8 десатурази, описаних в даному описі, замінюють на функціональний домен альтернативного гена десатурази, що, таким чином, приводить до утворення нового білка. В даному описі "домен" або "функціональний домен" позначає послідовність(ості) нуклеїнових кислот, здатні викликати біологічну реакцію в рослинах. Способи продукування різноманітних омега-3 та/або омега-6 жирних кислот Очікується, що введення химерних генів, які кодують дельта-8 десатуразу, описаних в даному описі (тобто, E389D8, E1594D8, TpomD8 або інші мутантні ферменти, оптимізовані за кодонами ферменти або їх гомологи), під контролем відповідних промоторів буде приводити до збільшення продукування ДГЛК та/або ЕТК в трансформованому організмі-хазяїні, відповідно. Таким чином, даний винахід включає спосіб прямого продукування ПНЖК що включає обробку субстрату жирних кислот (тобто, ЕДК та/або ЕТрК) десатуразними ферментами, описаними в даному описі (наприклад, E389D8, E1594D8, TpomD8), таким чином, що субстрат перетворюється на бажаний продукт у вигляді жирних кислот (тобто, ДГЛК та/або ЕТК). Більш конкретно, об'єктом даного винаходу є запропонувати спосіб ля продукування ДГЛК в клітині-хазяїні (наприклад, олеогені дріжджі, соя), де клітина-хазяїн включає: (a) рекомбінантний конструкт, що кодує поліпептид дельта-8 десатурази вибраний з групи, що складається з SEQ ID NO: 47, SEQ ID NO: 49 і SEQ ID NO: 57; і, (b) джерело ЕДК; де клітину-хазяїн вирощують в таких умовах, що експресується дельта-8 десатураза, і ЕДК перетворюється на ДГЛК, і де ДГЛК необов'язково добувають. В альтернативних варіантах даного винаходу, дельта-8 десатурази може бути використана як фермент для перетворення ЕТрК до ЕТК. Відповідно, у винаході пропонується спосіб продукування ЕТК, де клітина-хазяїн включає: (a) рекомбінантний конструкт, що кодує поліпептид дельта-8 десатурази вибраний з групи, що складається з SEQ ID NO: 47, SEQ ID NO: 49 і SEQ ID NO: 57; і, (b) джерело ЕТрК; де клітину-хазяїн вирощують в таких умовах, що експресується дельта-8 десатураза, і ЕТрК перетворюється на ЕТК, і де ЕТК необов'язково добувають. Альтернативно, кожен ген дельта-8 десатурази і відповідний ферментний продукт, описаний в даному описі, може застосовуватися непрямим чином для продукування різноманітних омега6 і омега-3 ПНЖК, в тому числі, наприклад, ДГЛК, ЕТК, АРК, ЕПК, ДПК та/або ДГК (фіг. 6; див. PCT публікацію WO 2004/101757). Непряме продукування омега-3/омега-6 ПНЖК відбувається, коли субстрат жирних кислот перетворюється непрямим чином на бажаний продукт у вигляді жирних кислот, через засоби проміжної стадії(й) або проміжної сполуки(ук) шляху. Таким чином, передбачається, що дельта-8 десатурази, описані в даному винаході (тобто, E389D8, E1594D8, TpomD8, або інші мутантні ферменти, оптимізовані за кодонами ферменти або їх гомологи), можуть експресуватися в сполученні з додатковими генами, що кодують ферменти біосинтетичного шляху ПНЖК (наприклад, дельта-6 десатурази, C18/20 елонгази, дельта-17 десатурази, дельта-8 десатурази, дельта-15 десатурази, дельта-9 десатурази, дельта-12 десатурази, C14/16 елонгази, C16/18 елонгази, дельта-9 елонгази, дельта-5 десатурази, дельта-4 десатурази, C20/22 елонгази), для одержання вищих рівнів продукування омега-3/омега-6 жирних кислот з довгим ланцюгом (наприклад, АРК, ЕПК, ДПК і ДГК). У переважних варіантах, дельта-8 десатурази за даним винаходом будуть мінімально експресуватися в сполученні з дельта-9 елонгазою (наприклад, дельта-9 елонгаза, представлена в SEQ ID NO: 75, або оптимізована за кодонами дельта-9 елонгаза, 23 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 представлена в SEQ ID NO: 110). Однак, включення конкретних генів до конкретної касети експресії, буде залежати від клітини-хазяїна (і його профілю ПНЖК та/або профілю десатурази/елонгази), придатності субстрату і бажаного кінцевого продукту(ів). Іноді може бути бажаним мінімізувати утворення побічного продукту у вигляді жирних кислот. Відносна кількість побічного продукту у вигляді жирних кислот може бути зменшена шляхом збільшення загальної активності дельта-8 десатурази. Один з підходів до мінімізації кількості побічного продукту у вигляді жирних кислот полягає в експресії більш ніж однієї дельта-8 десатурази (тобто, такої ж або іншої дельта-8 десатурази). Наприклад, присутність скіадонової кислоти (СКІ) та/або юніперової кислоти (ЮНП) [звичайно знаходяться в ліпідах насіння голосім'яних (Wolff et al., Lipids 35(1):1-22 (2000)), наприклад, представників родини Pinaceae (сосна)], може вважалася побічним продуктом у вигляді жирних кислот шляху дельта-6 десатурази/дельта-6 елонгази або шляху дельта-9 елонгази/дельта-8 десатурази. Хоча вважається, що зазначені жирні кислоти самі по собі володіють різноманітними оздоровчими властивостями (Nakane et al., Biol. Pharm. Bull. 23: 758-761 (2000)), їх присутності як побічного продукту у вигляді жирних кислот в сконструйованому шляху ПНЖК, наприклад, в урожаї олійної культури, може бути небажаною, в залежності від застосування. Термін "шлях дельта-6 десатурази/дельта-6 елонгази" також позначає біосинтетичний шлях для продукуванням ПНЖК з довгим ланцюгом. Даний шлях, як мінімумі, включає дельта-6 десатуразу і дельта-6 елонгазу, таким чином дозволяючи біосинтез ДГЛК та/або ЕТК з ЛК та AЛК, відповідно. При експресії іншої десатурази і елонгази, АРК, ЕПК, ДПК і ДГК також можуть бути синтезовані. Іноді, дельта-6 елонгаза може подовжувати жирні кислоти, окрім цільової жирної кислоти. Наприклад, дельта-6 елонгази загалом перетворюють ГЛК на ДГЛК, але деякі дельта-6 елонгази можуть також перетворювати нецільові субстрати, наприклад, ЛК або AЛК до ЕДК або ЕТрК, відповідно. У шляху дельта-6 десатурази/дельта-6 елонгази, ЕДК і ЕТрК розглядаються як "побічний продукт у вигляді жирних кислот", як визначено нижче. Додавання дельта-8 десатурази до шляху дельта-6 десатурази/дельта-6 елонгази запропонувало б засоби зворотного перетворення "побічного продукту у вигляді жирних кислот" ЕДК і ЕТрК на "проміжні жирні кислоти" (як визначено нижче) ДГЛК і ЕТК, відповідно. Системи експресії в рослинах, касети і вектори, трансформація В одному варіанті даний винахід стосується рекомбінантного конструкту, що включає будьякий з полінуклеотидів дельта-8 десатурази за винаходом, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, придатною для експресії в рослині. Промотор являє собою послідовність ДНК, яка спрямовує клітинний апарат рослини на продукування РНК з суміжної кодуючої послідовності в напрямку 5’-3’ (в напрямку 3’) по відношенню до промотору. Ділянка промотору впливає на швидкість, стадію розвитку і тип клітини, в якій виготовляється РНК транскрипт гена. РНК транскрипт обробляється з одержанням мРНК, яка служить шаблоном для трансляції послідовності РНК в послідовність амінокислот кодованого поліпептиду. Нетрансльована 5’ лідерна послідовність представляє собою ділянку мРНК, розташована в напрямку 3’-5’ по відношенню до ділянки, що кодує білок, яка може відігравати роль в ініціації і трансляції мРНК. 3’ сигнал термінації транскрипції/поліаденілювання представляє собою нетрансльовану ділянку в напрямку 5’-3’ по відношенню до ділянки, що кодує білок, функція якої в клітині рослини полягає у спричиненні термінації транскрипта РНК і додаванні поліаденілатних нуклеотидів до 3’ кінця РНК. Походження промотору, вибраного для контролю експресії послідовності, що кодує дельта-8 десатуразу, не є критичних до тих пір, поки промотор володіє достатньою транскрипційною активністю, щоб забезпечити експресію за винаходом трансльованої мРНК для цільових фрагментів нуклеїнових кислот в бажаній тканині хазяїна в потрібний час. Гетерологічні або негетерологічні (тобто, ендогенні) промотори можуть застосовуватися для практики винаходу. Наприклад, придатні промотори включають, не обмежуючись ними: субодиницю альфа прайм промотору бета конгліциніну, промотор інгібітора 3 трипсину Куніца (Kunitz), анексиновий промотор, промотор гліциніну Gy1, субодиницю бета промотору бета конгліциніну, промотор P34/Gly Bd m 30K, альбуміновий промотор, промотор A1 Leg і промотор A2 Leg. Анексиновий або P34 промотор описаний в PCT публікації WO 2004/071178 (опублікована 26 серпня 2004 р.). Рівень активності анексинового промотору є порівнянним з рівнем активності багатьох з відомих потужних промоторів, таких як: (1) промотор CaMV 35S (Atanassova et al., Plant Mol. Biol. 37:275-285 (1998); Battraw and Hall, Plant Mol. Biol. 15:527-538 (1990); Holtorf et al., Plant Mol. Biol. 29:637-646 (1995); Jefferson et al., EMBO J. 6:3901-3907 (1987); Wilmink et al., Plant Mol. Biol. 28:949-955 (1995)); (2) олеозинові промотори Arabidopsis (Plant et al., Plant Mol. Biol. 25:193-205 (1994); Li, Texas A&M University Ph.D. dissertation, pp. 107128 (1997)); (3) промотори білка подовження убіхітину Arabidopsis (Callis et al., J Biol. Chem. 24 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 265(21):12486-93 (1990)); (4) промотори гена убіхітину помідора (Rollfinke et al., Gene. 211(2):267-76 (1998)); (5) промотор соєвого білка теплового удару (Schoffl et al., Mol Gen Genet. 217(2-3):246-53 (1989)); і, (6) промотор гена гістона H3 маїсу (Atanassova et al., Plant Mol Biol. 37(2):275-85 (1989)). Інша корисна ознака анексинового промотору полягає в його профілі експресії в насінні, що розвивається. Анексиновий промотор виявляє найбільшу активність на ранніх стадіях розвитку зерен (до 10 днів після запилення) і є істотною мірою латентним на більш пізніх стадіях. Профіль експресії анексинового промотору відмінний від профілю експресії багатьох специфічних для насіння промоторів, наприклад, промоторів запасного білка, які часто демонструють найвищу активність на більш пізніх стадіях розвитку (Chen et al., Dev. Genet. 10:112-122 (1989); Ellerstrom et al., Plant Mol. Biol. 32:1019-1027 (1996); Keddie et al., Plant Mol. Biol. 24:327-340 (1994); Plant et al. [вище]; Li, [вище]). Профіль експресії анексинового промотору більш традиційний, але залишається відмінним від інших відомих промоторів, специфічних для насіння. Таким чином, анексиновий промотору буде дуже привабливим кандидатом, якщо надмірна експресія або пригнічення, гена в ембріонах є бажаними га ранніх стадіях розвитку. Наприклад, може бути бажаною надмірна експресія гена, що регулюючий ранній розвиток ембріону, або гена, залученого до метаболізму перед стадіями дозрівання зерна. Після ідентифікації відповідного промотору, придатного для експресії конкретної кодуючої послідовності дельта-8 десатурази, забезпечують функціональне зв'язування промотору в смисловій орієнтації, із застосуванням звичних засобів, добре відомі фахівцям в даній галузі. Стандартна рекомбінантна ДНК і молекулярні методи клонування, використані в даному описі, добре відома в даній галузі і описані більш детально в Sambrook, J. et al., в Molecular Cloning: A Laboratory Manual; 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York, 1989 (hereinafter "Sambrook et al., 1989") або Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A. and Struhl, K., Eds.; в Current Protocols in Molecular Biology; John Wiley and Sons: New York, 1990 (hereinafter "Ausubel et al., 1990"). Після виготовлення, рекомбінантний конструкт може бути введений у вибрану рослинну клітину способами, добре відомими середньому фахівцю в даній галузі (наприклад, трансфекція, трансформація та електропорація). Клітини рослин олійних культур є переважними рослинними клітинами. Перетворену рослинну клітину далі культивують і вирощують в придатних умовах, що дозволяють експресію ПНЖК з довгим ланцюгом, яку далі необов'язково добувають та очищують. Рекомбінантні конструкти за винаходом можуть бути введені в одну рослинну клітину; або, альтернативно, кожний конструкт може бути введений в окрему рослинну клітину. Експресія в рослинній клітині може бути здійснена у стабільній або тимчасовій манері, як описано вище. Бажані ПНЖК з довгим ланцюгом можна експресувати в зерні. Також в межах контексту даного винаходу знаходяться зерна або частини рослини, одержані з таких трансформованих рослин. Частини рослини включають диференційовані і недиференційовані тканини, в тому числі, не обмежуючись ними, наступні: коріння, стовбури, бруньки, листя, пилок, зерна, пухлинні тканини і різні форми клітин та культури (наприклад, одиничні клітини, протопласти, ембріони і тканина калюсу). Тканина рослини може знаходитися в рослині або в органі рослини, тканині або культурі клітин. Термін "орган рослини" позначає тканину або групу тканин рослини, які складають морфологічно і функціонально відмінну частину рослини. Термін "геном" позначає наступне: (1) повний комплемент генетичного матеріалу (гени і некодуючі послідовності), що присутній в кожній клітині організму або вірусу або органели; та/або (2) комплект хромосом, що успадковується як набір від одного батька (гаплоїдний). Таким чином, даний винахід також стосується способу трансформації клітини, який включає трансформацію клітини за допомогою рекомбінантного конструкта за винаходом і селекції таких клітин, трансформованих за допомогою рекомбінантного конструкта за п. 8. Також представляє інтерес спосіб одержання трансформованої рослини, який включає трансформацію рослинної клітини за допомогою полінуклеотидів дельта-8 десатурази за даним винаходом і вирощування рослини з трансформованої рослинної клітини. Способи трансформації дводольних рослин (головним чином, із застосуванням Agrobacterium tumefaciens) та одержання трансгенних рослин опубліковані, серед іншого, для: бавовни (патент США 5,004,863; патент США 5,159,135); сої (патент США 5,569,834; патент США 5,416,011); Brassica (патент США 5,463,174); арахісу (Cheng et al. Plant Cell Rep. 15:653-657 (1996); McKently et al. Plant Cell Rep. 14:699-703 (1995)); папайї (Ling, K. et al. 25 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Bio/technology 9:752-758 (1991)) і гороху (Grant et al. Plant Cell Rep. 15:254-258 (1995)). Огляду інших традиційних способів трансформації рослини див. у Newell, C.A. (Mol. Biotechnol. 16:53-65 (2000)). В одному з вказаних способів трансформації використовують Agrobacterium rhizogenes (Tepfler, M. and Casse-Delbart, F. Microbiol. Sci. 4:24-28 (1987)). Описане перетворення сої із застосуванням безпосередньої доставки ДНК за допомогою ПЕГ злиття (PCT публікація WO 92/17598), електропорації (Chowrira, G.M. et al., Mol. Biotechnol. 3:17-23 (1995); Christou, P. et al., Proc. Natl. Acad. Sci. U.S.A. 84:3962-3966 (1987)), мікроін'єкції і бомбардування частинками (McCabe, D.E. et. al., Bio/Technology 6:923 (1988); Christou et al., Plant Physiol. 87:671-674 (1988)). Існують численні способи регенерації рослин з рослинної тканини. Конкретний метод регенерації буде залежати від вихідної рослинної тканини і конкретних видів рослин, які підлягають регенерації. Регенерація, розвиток і культивування рослин з трансформантів у вигляді єдиного протопласту рослини або з різних трансформованих експлантатів добре відомі в даній галузі (Weissbach and Weissbach, In: Methods for Plant Molecular Biology, (Eds.), Academic: San Diego, CA (1988)). Такий процес регенерації та вирощування звичайно включає стадії селекції трансформованих клітин і культивування таких індивідуалізованих клітин через звичайні стадії ембріонального розвитку, в тому числі через стадію саджанця, що вкоренився. Трансгенні ембріони і зерна регенерують подібним чином. Одержані трансгенні пагони, що вкоренилися далі висаджують у відповідне середовище для росту рослин, таке як грунт. Переважно, регенеровані рослини самозапилюються, щоб забезпечити гомозиготні трансгенні рослини. В іншому варіанті, пилок, одержаний з регенерованих рослин переносять на вирощені з насіння рослини важливих з агрономічної точки зору ліній. З іншого боку, пилок рослин з таких важливих ліній застосовують для запилення регенерованих рослин. Трансгенну рослину за даним винаходом, що містить бажаний поліпептид, культивують із застосуванням способів, добре відомих фахівцю в даній галузі. На додаток до обговорених вище методик, практикуючі фахівці знайомі із стандартними матеріалами з джерел, де описані конкретні умови і методики: конструювання, маніпулювання і виділення макромолекул (наприклад, молекули ДНК, плазміди і т. п.); генерації рекомбінантних фрагментів ДНК і рекомбінантних конструктів експресії; а також скринінгу та виділення клонів. Див., наприклад: Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor: NY (1989); Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor: NY (1995); Birren et al., Genome Analysis: Detecting Genes, Vol.1, Cold Spring Harbor: NY (1998); Birren et al., Genome Analysis: Analyzing DNA, Vol.2, Cold Spring Harbor: NY (1998); Plant Molecular Biology: A Laboratory Manual, eds. Clark, Springer: NY (1997). Приклади рослин олійної культури включають, не обмежуючись ними: сою, види Brassica, соняшник, маїс, бавовну, льон і сафлор. Приклади ПНЖК, що містять як мінімум 20 атомів вуглецю і п'ять або більше подвійних вуглець-вуглецевих зв'язків включають, не обмежуючись ними, омега-3 жирні кислоти, наприклад, ЕПК, ДПК і ДГК. Зерна, одержані з таких рослин, також знаходяться в межах контексту даного винаходу, а також як олія, одержана з таких зерен. Таким чином, в одному варіанті даний винахід стосується рослини олійної культури, яка включає: (a) перший рекомбінантний конструкт ДНК, що містить виділений полінуклеотид, який кодує поліпептид дельта-8 десатурази, функціонально зв'язаний як мінімум з однією регуляторною послідовністю; і, (b) як мінімум один додатковий рекомбінантний конструкт ДНК, що містить виділений полінуклеотид, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, який кодує поліпептид, вибраний з групи, що складається з дельта-4 десатурази, дельта-5 десатурази, дельта-6 десатурази, дельта-8 десатурази, дельта-9 десатурази, дельта-9 елонгази, дельта-12 десатурази, дельта-15 десатурази, дельта-17 десатурази, C14/16 елонгази, C16/18 елонгази, C18/20 елонгази і C20/22 елонгази. Такі додаткові десатурази обговорюється, наприклад, в патентах США 6,075,183, 5,968,809, 6,136,574, 5,972,664, 6,051,754, 6,410,288 і PCT публікаціях WO 98/46763, WO 98/46764, WO 00/12720 і WO 00/40705. Вибір комбінації касет для використання частково залежить від профілю ПНЖК та/або профілю десатурази/елонгази клітин рослини олійної культури, які підлягають трансформації, та від ПНЖК з довгим ланцюгом, який повинні бути експресовані. В іншому аспекті, даний винахід стосується способу одержання ПНЖК з довгим ланцюгом у рослинній клітині, який включає: (a) трансформацію клітини за допомогою рекомбінантного конструкта за винаходом; і, (b)селекцію трансформованих клітин, які продукують ПНЖК з довгим ланцюгом. 26 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Ще в іншому аспекті, даний винахід стосується способу одержання як мінімум однієї ПНЖК у клітині сої, який включає: (a) трансформацію соєвої клітини за допомогою першого рекомбінантного конструкта ДНК, що містить: (i) виділений полінуклеотид, який кодує поліпептид дельта-8 десатурази, функціонально зв'язаний як мінімум з однією регуляторною послідовністю; і, (ii) як мінімум один додатковий рекомбінантний конструкт ДНК, що містить виділений полінуклеотид, функціонально зв'язаний як мінімум з однією регуляторною послідовністю, який кодує поліпептид, вибраний з групи, що складається з дельта-4 десатурази, дельта-5 десатурази, дельта-6 десатурази, дельта-8 десатурази, дельта-9 десатурази, дельта-9 елонгази, дельта-12 десатурази, дельта-15 десатурази, дельта-17 десатурази, C14/16 елонгази, C16/18 елонгази, C18/20 елонгази і C20/22 елонгази; (b) регенерацію рослини сої з трансформованої клітини із стадії (a); і, (c) селекцію зерен, одержаних з рослин із стадії (b), що містять змінений рівень ПНЖК в порівнянні з рівнем в зернах, одержаних з нетрансформованої соєвої рослини. В інших переважних варіантах, як мінімум один додатковий рекомбінантний конструкт ДНК кодує поліпептид, який володіє активністю дельта-9 елонгази, наприклад, дельта-9 елонгази, що виділена або походить з Isochrysis galbana (інвентарний номер GenBank AF390174; IgD9e), представленої в SEQ ID NO: 76, або дельта-9 елонгази, що виділена або походить з Euglena gracilis, представленої в SEQ ID NO: 75. Мікробні системи експресії, касети і вектори, трансформація Гени дельта-8 елонгази і генні продукти, описані в даному винаході (тобто, E389D8, E1594D8, TpomD8, або інші мутантні ферменти, оптимізовані за кодонами ферменти або їх гомологи), також можуть бути продуковані в гетерологічних мікробних клітинах-хазяях, особливо в клітинах олеогених дріжджів (наприклад, Yarrowia lipolytica). Мікробні системи експресії і вектори експресії, що містять регуляторні послідовності, які спрямовують високі рівні експресії чужорідних білків, добре відомі фахівцям в даній галузі. Будьякі з них можуть застосовуватися для конструювання химерних генів для продукування будьякого з генних продуктів необхідних послідовностей. Такі химерні гени далі можуть бути введені у відповідні мікроорганізми шляхом трансформації для забезпечення високих рівнів експресії кодованих ферментів. Вектори або касети ДНК, придатні для трансформації придатних мікробних клітин-хазяїв, добре відомі в даній галузі. Конкретний вибір послідовностей, присутніх в конструкті, залежить від бажаних продуктів експресії (вище), природи клітини-хазяїна і запропонованих засобів відокремлення трансформованих клітин від нетрансформованих клітин. Однак, звичайно вектор або касета містить послідовності, що спрямовують транскрипцію і трансляцію відповідного гена(ів), селекційний маркер і послідовності, що дозволяють автономну реплікацію або інтеграцію в хромосому. Придатні вектори включають ділянку 5' гена, яка контролює ініціацію транскрипції (наприклад, промотор), і ділянку 3' фрагмента ДНК, яка контролює термінацію транскрипції (тобто, термінатор). Найбільш переважно, обидві контролюючі ділянки походять з генів трансформованої мікробної клітини-хазяїна, хоча слід розуміти, що такі контрольні ділянки не повинні обов'язково бути одержані з генів, природних для конкретних видів, вибраних як хазяїн продукування. Ділянки контролю ініціації або промотори, придатні для управління експресією необхідних ВРЗ дельта-8 десатурази в цільовій мікробній клітині-хазяїні, численні і знайомі фахівцям в даній галузі. Фактично будь-який промотор, здатний до спрямування експресії таких генів у відібраній клітині-хазяїні, є придатним для цілей даного винаходу. Експресія в мікробній клітиніхазяїні може здійснюватися в режимі стабільної або тимчасової експресії. Тимчасова експресія може бути забезпечена індукуванням активності промотору, який піддається регулюванню, функціонально зв'язаного з цільовим геном. Стабільна експресія може бути досягнута застосуванням конститутивного промотору, функціонально зв'язаного з цільовим геном. Як приклад, якщо клітина-хазяїн відноситься до дріжджів, забезпечуються транскрипційні і трансляційні ділянки, функціональні в клітинах дріжджів, особливо з видів хазяїв (наприклад, див. PCT публікації WO 2004/101757 і WO 2006/052870 щодо переважних регуляторних ділянок для ініціації транскрипції з метою застосування в Yarrowia lipolytica). Будь-яка з цілого ряду регуляторних послідовностей може застосовуватися, в залежності від того, конститутивна чи індукована транскрипція є бажаною, ефективності промотору з точки зору експресії цільової ВРЗ, легкості конструювання і т. п. 27 UA 102059 C2 5 10 15 20 25 30 35 40 45 50 55 60 Виявлено, що нуклеотидні послідовності, які оточують кодон ініціації трансляції "ATG" впливають на експресію в клітинах дріжджів. Якщо цільовий поліпептид слабко експресується в дріжджах, нуклеотидні послідовності екзогенних генів можуть бути модифіковані таким чином, щоб включати ефективну послідовність ініціації трансляції дріжджів для одержання оптимальної експресії гена. Для експресії в дріжджах, це може бути здійснено сайт-спрямованим мутагенезом гена, що недостатньо експресується, шляхом злиття його в рамці з ендогенним геном дріжджів, переважно гена з високою експресією. Альтернативно, можна визначити консенсусну послідовність ініціації трансляції в хазяїні і сконструювати дану послідовність в гетерологічних генах для їх оптимальної експресії в цільовому хазяїні. Ділянка термінації може походити від 3' ділянки гена, з якого одержана ділянка ініціації, або з іншого гена. Велика кількість ділянок термінації відома і задовільно функціонує в різноманітних хазяях (при використанні в такому ж та інших родах і видах, в порівннянні з тим, з якого вони були одержані). Ділянку термінації звичайно вибирають більшою мірою за традицією, ніж через будь-яку конкретну властивість. Переважно, якщо мікробний хазяїн представляє собою дріжджову клітину, ділянка термінації походить з гена дріжджів (особливо Saccharomyces, Schizosaccharomyces, Candida, Yarrowia або Kluyveromyces). Також відомо, що 3'-ділянки ссавцевих генів, що кодують -інтерферон та -2 інтерферон функціонують в дріжджах. Ділянки контролю термінації можуть також походити з різних генів, природних для переважних хазяїв. Необов'язково, сайт термінації може бути непотрібним; однак, найбільш переважно, він включений. Без наміру обмежувати винахід, ділянки термінації, придатні для розкриття в даному описі, включають: ~100 пари основ 3’ ділянки позаклітинної протеази Yarrowia lipolytica (XPR; інвентарний номер GenBank M17741); термінатори ацил-КоА оксидази (Aco3: інвентарні номери GenBank AJ001301 і CAA04661; Pox3: інвентарний номер GenBank XP_503244); термінатор Pex20 (інвентарний номер GenBank AF054613); термінатор Pex16 (інвентарний номер GenBank U75433); термінатор Lip1 (інвентарний номер GenBank Z50020); термінатор Lip2 (інвентарний номер GenBank AJ012632); і термінатор 3-оксоацил-КоА тіолази (OCT; інвентарний номер GenBank X69988). Як відомо фахівцю в даній галузі, сама тільки вставка гена у вектор клонування не гарантує успішної експресії на необхідному рівні. У відповідь на потребу у високій швидкості експресії, створені численні спеціалізовані вектори експресії шляхом маніпулювання цілим рядом різноманітних генетичних елементів, які контролюють аспекти транскрипції, трансляції, стабільності білка, обмеження кисню і секреції з мікробної клітини-хазяїна. Більш конкретно, деякі з молекулярних ознак, якими маніпулюють для контролю експресії гена, включають: (1) природу відповідних послідовностей промотору і термінатора транскрипції; (2) кількість копій клонованого гена і статус гена (походить з плазміди чи інкорпорований в геном клітини-хазяїна); (3) кінцеву локалізацію в клітині розташування синтезованого чужорідного білка; (4) ефективність трансляції і правильне складання білка в організмі-хазяїні; (5) внутрішню стабільність мРНК і білка клонованого гена в межах клітини-хазяїна; і (6) використання кодону в межах клонованого гена, таким чином, що його частота наближається до частота переважного використання кодону клітиною-хазяїном. Кожний з цих типів модифікацій включений до даного винаходу, оскільки має на меті подальше оптимізацію експресії дельта-8 десатурази, описаної в даному винаході. Після одержання ДНК (наприклад, химерний ген, що включає промотор, ВРЗ і термінатор), що кодує поліпептид, придатний для експресії у відповідній мікробній клітині-хазяїні (наприклад, олеогені дріжджі), її вміщують у плазмідний вектор, здатний до автономної реплікації в клітиніхазяїні, або безпосередньо інтегрують в геном клітини-хазяїна. Інтеграція касет експресії може відбуватися випадковим чином в межах генома хазяїна або може бути цілеспрямованою шляхом застосування конструктів, що містять ділянки гомології з геномом хазяїна, достатньої для цільової рекомбінації в межах локусу хазяїна. Якщо конструкти спрямовані на ендогенний локус, всі або деякі з транскрипційних і трансляційних регуляторних ділянок можуть бути забезпечені ендогенним локусом. В даному винаході, переважний спосіб експресії генів в Yarrowia lipolytica полягає в інтеграції лінійної ДНК в геном хазяїна; і інтеграція в множинні місця розташування в межах генома може бути особливо придатною, якщо бажаною є високий рівень експресії генів [наприклад, в локусі Ura3 (інвентарний номер GenBank AJ306421), локусі гена Leu2 (інвентарний номер GenBank AF260230), гені Lys5 (інвентарний номер GenBank M34929), локусі гена Aco2 (інвентарний номер GenBank AJ001300), локусі гена Pox3 (Pox3: інвентарний номер GenBank XP_503244; або Aco3: інвентарний номер GenBank AJ001301), локусі гена дельта-12 десатурази (PCT публікація WO 2004/104167), локусі гена Lip1 (інвентарний номер GenBank Z50020) та/або локусі гена Lip2 (інвентарний номер GenBank AJ012632)]. 28
ДивитисяДодаткова інформація
Назва патенту англійськоюDelta-8-desaturase and it using for production of polyunsaturated fatty acids
Автори англійськоюDamude, Howard, Zhu, Quinn, Qun, Xue, Zhixiong
Назва патенту російськоюДельта-8 десатураза и ее применение для получения полиненасыщенных жирних кислот
Автори російськоюДамуд Ховард, Жу Квинн Кун, Су Жисонг
МПК / Мітки
МПК: C12N 15/82, A01H 5/10, C12N 9/02, C12N 15/53
Мітки: кислот, одержання, дельта-8, поліненасичених, десатураза, застосування, жирних
Код посилання
<a href="https://ua.patents.su/332-102059-delta-8-desaturaza-ta-zastosuvannya-dlya-oderzhannya-polinenasichenikh-zhirnikh-kislot.html" target="_blank" rel="follow" title="База патентів України">Дельта-8 десатураза та її застосування для одержання поліненасичених жирних кислот</a>
Застосування довголанцюжкових поліненасичених жирних кислот для одержання рідкої поживної суміші для годування дітей

Номер патенту: 89207
Опубліковано: 11.01.2010
Автори: Бем Гюнтер, Берманн Крістофер, Шталь Бернд, Гарссен Йохан
МПК: A23L 1/29, A61K 31/202, A61P 43/00
Мітки: дітей, годування, довголанцюжкових, кислот, поживної, одержання, застосування, суміші, жирних, поліненасичених, рідкої
Формула / Реферат:
1. Застосування довголанцюжкових поліненасичених жирних кислот для одержання рідкої поживної суміші для годування дітей, матері яких страждали під час вагітності на діабет І типу, діабет II типу, ожиріння або надлишкову вагу, причому вказана суміш містить n-3 фракцію поліненасичених жирних кислот, що містить щонайменше 0,1 ваг. % докозагексаєнової кислоти (DHA) від повної ваги ліпіду, щонайменше 0,01 ваг. % n-3 докозапентаєнової кислоти...
Спосіб одержання поліненасичених жирних кислот в трансгенних організмах

Номер патенту: 90455
Опубліковано: 11.05.2010
Автори: Бауер Йорг, Домерг Фредерік, Врінтен Патрісія, Цірпус Петра, Цанк Торстен, Кірш Єлєна, Меєр Астрід, Шперлінг Петра, Квіу Ксяо, Аббаді Аміне, Хайнц Ернст
МПК: C12N 15/82, C12N 9/10, C12N 9/02
Мітки: спосіб, трансгенних, кислот, жирних, поліненасичених, організмах, одержання
Формула / Реферат:
1. Спосіб одержання сполук загальної формули І в трансгенних організмах із вмістом принаймні 1 мас.% даних сполук від загального вмісту ліпідів трансгенного організму, який відрізняється тим, що включає наступні етапи: a) введення в організм принаймні однієї послідовності нуклеїнової кислоти, яка кодує Δ-9-елонгазну або Δ-6-десатуразну...
Спосіб оцінки порушень метаболізму поліненасичених жирних кислот ліпідів ліквору
Номер патенту: 22884
Опубліковано: 05.05.1998
Автори: Пічкур Ліонид Дмитрович, Брюзгіна Тетяна Семенівна, Цимбалюк Віталій Іванович, Яценко Валентин Порфір'євич
МПК: G01N 30/68, C07C 57/00, G01N 33/53
Мітки: жирних, поліненасичених, спосіб, кислот, порушень, метаболізму, ліквору, оцінки, ліпідів
Формула / Реферат:
Способ оценки нарушений метаболизма полиненасыщенных жирных кислот липидов ликвора путем газохроматографического анализа, отличающийся тем, что определяют содержание арахидоновой и линолевой кислот липидов ликвора и по коэффициенту соотношения между ними устанавливают нарушение метаболизма полиненасыщенных жирных кислот.
Спосіб отримання гідропероксидів поліненасичених жирних кислот

Номер патенту: 13716
Опубліковано: 25.04.1997
Автори: Кухар Валерій Павлович, Бутович Ігор Анатолійович, Цись Олена Валентинівна, Бридня Віта Петрівна
МПК: C12N 11/00, G01N 1/28, C12P 7/64
Мітки: спосіб, гідропероксидів, жирних, поліненасичених, отримання, кислот
Формула / Реферат:
Способ получения гидропероксидов полиненасыщенных жирных кислот, включающий окисление полиненасыщенных жирных кислот в присутствии лйпоксигеназы в качестве катализатора и последующее выделение целевого продукта, отличающийся тем, что, с целью увеличения выхода целевого продукта и упрощения способа, окисление проводят в непрерывном режиме путем пропускания водного раствора полинепасыщсн-ных жирных кислот с концентрацией 0,01-0,25 мМ/л...
Жирова композиція з низьким вмістом транс-жирних кислот, спосіб її одержання та застосування для кондитерських і хлібобулочних виробів

Номер патенту: 78990
Опубліковано: 10.05.2007
Автори: ван Дік Марій, Саей Карен, Клеєневерк Бернард
МПК: A23D 9/02, A23G 3/00, C11C 3/00, A21D 13/08, A23D 9/00, A23G 1/00, A23G 3/34, A21D 2/16, A23G 1/38, A21D 13/00
Мітки: одержання, застосування, композиція, низьким, кондитерських, транс-жирних, спосіб, хлібобулочних, вмістом, кислот, жирова, виробів
Формула / Реферат:
1. Спосіб одержання жирової композиції, призначеної для застосування як нетемперованого кондитерського жиру або як жиру для виробництва хлібобулочних виробів, який відрізняється тим, що піддають вихідну жирову композицію, яка містить пальмову олію або фракцію пальмової олії і має такий склад:(1) гліцеридний склад ізвмістом S2U від 47 до 75 мас. %,вмістом SU2 + U3 менше 40 мас. %,вмістом S3 від 1 до 15 мас. %...
Попередній патент: Свердловинний фільтр і спосіб його експлуатації
Наступний патент: Загородження
Випадковий патент: Спосіб ослаблення тропічних циклонів (ураганів)