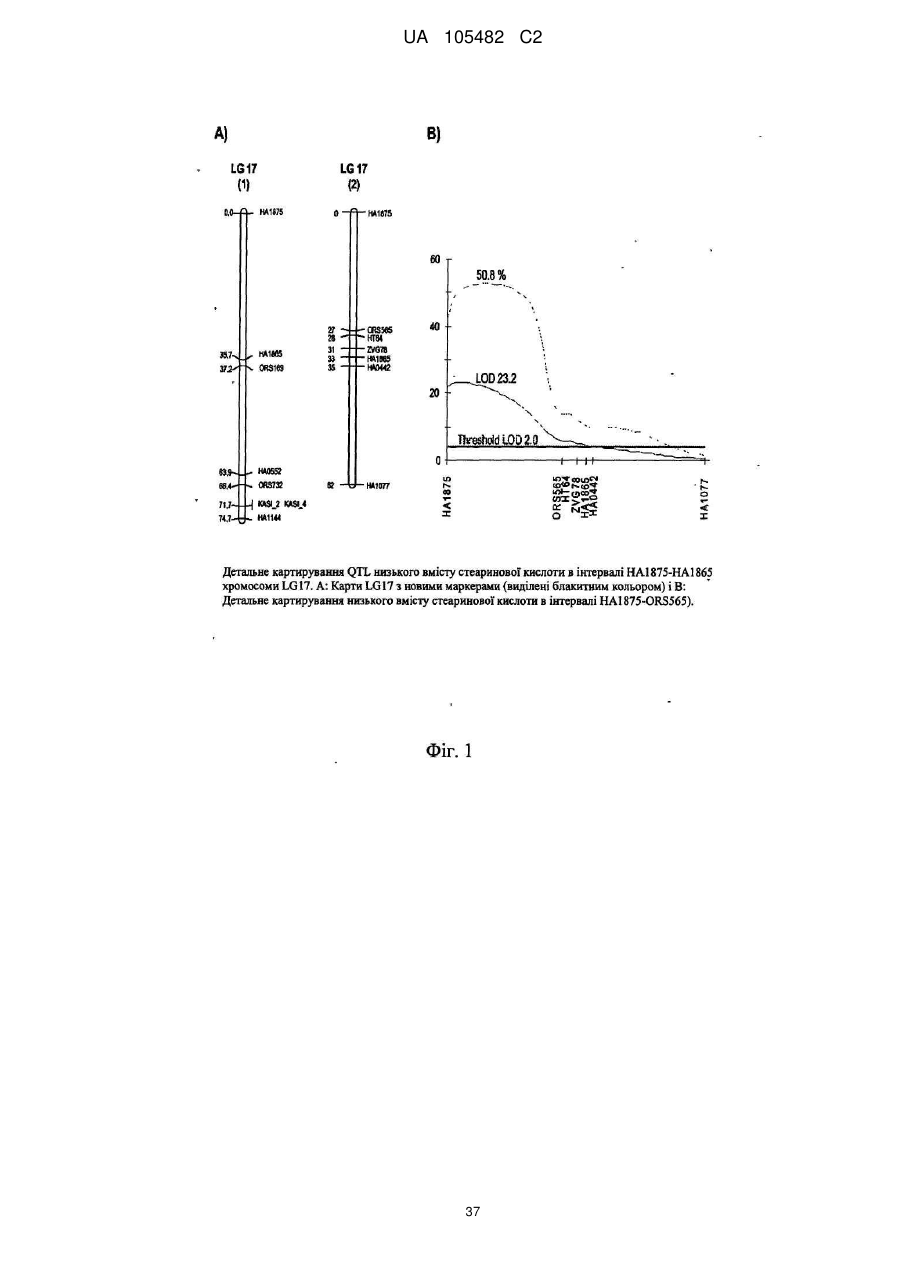
Соняшник з низьким вмістом насичених жирів
Номер патенту: 105482
Опубліковано: 26.05.2014
Автори: Кел Чарльз Джеймс, Гердес Джеймс Тодд, Еріксон Анджела Лі, Бенсон Роберт Мартін




































Формула / Реферат
1. Насіння соняшника, в якому композиція жирних кислот олії в насінні включає приблизно 3,0 % або менше загальної кількості взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0), причому соняшник був генетично модифікований шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичених жирів.
2. Насіння соняшника за п. 1, в якому вміст олії в насінні містить взяті разом пальмітинову кислоту (16:0) і стеаринову кислоту (18:0) в кількості приблизно або менше 2,8 %.
3. Рослина соняшника, або її частина, одержана вирощуванням насіння за п. 1.
4. Соняшник, який відрізняється тим, що має олію насіння, яка містить приблизно 3,0 % або менше загального вмісту взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0), причому соняшник генетично модифікований шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичених жирів.
5. Соняшник за п. 4, в якому олія насіння включає загальний вміст взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно або менше 2,8 %.
6. Соняшник за п. 4, в якому соняшник додатково має резистентність до гербіциду, вибраного з групи, що складається з імідазолінону, сульфонілсечовини, гліфосату, глюфозинату, L-фосфінотрицину, триазину, Clearfield, Dicamba, 2,4-D і бензонітрилу.
7. Насіння щонайменше одного соняшника, генетично модифікованого шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичених жирів, що містить приблизно 3,0 % або менше загального вмісту взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0).
8. Насіння за п. 7, в якому олія насіння включає загальний вміст взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно або менше 2,8 %.
9. Валова проба олії соняшника, екстрагованої з насіння соняшника, генетично модифікованого шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичених жирів, причому вказана олія соняшника має профіль жирних кислот, який включає приблизно 3,0 % або менше загального вмісту взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0).
10. Валова проба олії соняшника, екстрагованої з насіння соняшника за п. 9, в якій профіль жирних кислот включає загальний вміст взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно або менше 2,8 %.
11. Олія з насіння соняшника, генетично модифікованого шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичених жирів, що має профіль жирних кислот, який включає приблизно 3,0 % або менше загального вмісту взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0).
12. Олія з насіння соняшника за п. 11, в якій профіль жирних кислот негідрогенізованої олії включає загальний вміст взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно або менше 2,8 %.
13. Олія в насінні соняшника, генетично модифікованого шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем насичення, що має профіль вмісту жирних кислот, який включає приблизно 3,0 % або менше загального вмісту взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0).
14. Олія за п. 13 для харчового застосування.
15. Харчовий продукт, що містить олію за п. 13.
16. Композиція рослинної олії, що містить олію за п. 13.
17. Насіння соняшника за п. 1, в якому вміст олії в насінні включає приблизно 15 % або більше вмісту лінолевої кислоти (18:2).
18. Насіння соняшника за п. 17, в якому загальний вміст лінолевої кислоти (18:2) складає більше ніж приблизно 20 %.
19. Насіння соняшника за п. 17, в якому загальний вміст лінолевої кислоти (18:2) складає більше ніж приблизно 74 %.
20. Рослина соняшника, або її частина, одержана вирощуванням насіння за п. 17.
21. Рослина соняшника за п. 4, яка має рослинну олію, що включає приблизно 15 % або більше вмісту лінолевої кислоти (18:2).
22. Рослина соняшника за п. 21, в якій олія насіння включає загальний вміст лінолевої кислоти (18:2) приблизно або більше 20 %.
23. Валова проба олії соняшника, екстрагованої з насіння соняшника за п. 9, причому вказана олія соняшника має профіль жирних кислот, який включає приблизно 15 % або більше вмісту лінолевої кислоти (18:2).
24. Валова проба олії соняшника, екстрагованої з насіння соняшника, за п. 23, причому профіль жирних кислот олії включає загальний вміст лінолевої кислоти (18:2) приблизно або більше 20%.
25. Олія, екстрагована з насіння соняшника за п. 11, де вказана олія має вміст жирних кислот, який включає приблизно 15 % або більше вмісту лінолевої кислоти (18:2).
26. Олія з насіння соняшника за п. 25, причому профіль жирних кислот олії включає загальний вміст лінолевої кислоти (18:2) приблизно або більше 20 %.
27. Олія в насінні соняшника за п. 13, причому вказана олія має профіль жирних кислот, який включає приблизно 15 % або більше вмісту лінолевої кислоти (18:2).
28. Олія за п. 27 для харчового застосування.
29. Харчовий продукт, що містить олію за п. 27.
30. Композиція рослинної олії, що містить олію за п. 27.
Текст
Реферат: Винахід належить до рослини соняшника та її насіння, в якому композиція жирних кислот олії включає приблизно 3,0 % або менше загальної кількості взятих разом пальмітинової кислоти (16:0) і стеаринової кислоти (18:0), причому соняшник генетично модифікований шляхом селекції рослин таким чином, що він включає зародкову плазму з низьким рівнем ненасичених жирів, і необов’язково з високим вмістом лінолевої кислоти. Також винахід належить до олії, харчового продукту та композиції рослинної олії, які отримані з такої рослини соняшника та її насіння. UA 105482 C2 (12) UA 105482 C2 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 ДОМАГАННЯ ПРІОРИТЕТУ По цій заявці вимагається пріоритет за датою подачі тимчасової заявки на патент США з серійним номером 61/015591, поданої 20 грудня 2007 року. ГАЛУЗЬ ТЕХНІКИ Даний винахід стосується нових і відмінних рослин соняшника, що дають насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, а також відповідних способів. Даний винахід додатково стосується не модифікованих генетично, немутованих рослин соняшника, що мають стійкість до гліфосату, і відповідним способам. ПЕРЕДУМОВИ ВИНАХОДУ Соняшник, що культивується, (Helianthus annuus L.) є основним світовим джерелом рослинної олії. У Сполучених Штатах Америки приблизно 4 мільйони акрів щорічно засіваються соняшником, переважно в Дакоті і Мінесоті. Дуже швидке збільшення кількості акрів, що засіваються соняшником, за останнє десятиріччя в Сполучених Штатах Америки частково є наслідком декількох важливих розробок в галузі селекції соняшника і поліпшення сортів. Однією з важливих розробок було відкриття цитоплазматичної чоловічої стерильності і генів для відновлення фертильності, яке дозволило одержання гібридних рослин соняшника. Одержані таким чином гібриди були введені в культуру на початку 1970-х років. Опис цитоплазматичної чоловічої стерильності (ЦМС) і генетичне відновлення фертильності у соняшника представлений в Fick, «Breeding and Genetics» в Sunflower Science and Technology 279-338 (J.F. Carter ed. 1978). Олія соняшника складається в основному з пальмітинової (16:0), стеаринової (18:0), олеїнової (18:1), лінолевої (18:2) і ліноленової (18:3) кислот. Хоча в рослинах присутні інші незвичайні жирні кислоти, пальмітинова, стеаринова, олеїнова, лінолева і ліноленова кислоти становлять приблизно 88% жирних кислот, присутніх в світовому виробництві рослинних олій. (J.L. Harwood, Plant Acyl Lipids: Structure, Distribution and Analysis, 4 Lipids: Structure and Function, P.K. Stumpf and E.E. Conn ed. (1988)). Пальмітинова і стеаринова кислоти є насиченими жирними кислотами, які, як було показано в деяких дослідженнях, вносять внесок в збільшення рівня плазматичного холестерину, причини коронарної хвороби серця. Згідно з останніми дослідженнями рослинні олії з високим вмістом ненасичених жирних кислот, таких як олеїнова і лінолева кислоти, може володіти здатністю знижувати рівень плазматичного холестерину. Насичені жирні кислоти також загалом мають більш високі точки плавлення відносно ненасичених жирних кислот з таким же числом атомів вуглецю, що вносить внесок в проблему стійкості до холоду продуктів харчування і може вносити внесок у відчуття воску або жиру у роті в ході їди. Також відомо, що харчові продукти, виготовлені з жирів і олій, що мають менше приблизно 3% насичених жирів будуть звичайно містити менше 0,5 грама насичених жирів на порцію, і в результаті при існуючих нормативах маркірування можуть бути маркіровані, як такі, «що не містять насичених жирів». Отже, з ряду причин бажано одержувати олію соняшника з низьким вмістом пальмітинової і стеаринової кислот і високим вмістом олеїнової або лінолевої кислот. Виведення будь-якої нової бажаної рослинної зародкової плазми складається з ряду стадій. Селекція рослин починається з аналізу і визначення проблем і недоліків зародкової плазми, що є в наявності, встановлення цілей програми і визначення конкретних задач схрещування. Наступною стадією є відбір зародкової плазми, що володіє ознаками, які задовольняють цілям програми. Метою є об'єднання в одному сорті поліпшеної комбінації бажаних ознак з батьківської зародкової плазми. Ці важливі ознаки можуть включати високий вихід насіння, резистентність до хвороб і комах, більш міцні стеблини і коріння, переносимість засухи і жари, і більш хорошу агротехнічну якість. Вибір способів селекції і відбору залежить від способу репродукції рослини, здібності успадкувати ознаку (ознаки), що поліпшується, і типу культивованого сорту, що використовується з комерційною метою (наприклад, F1-гібрид, чиста лінія і т.д.). Для високо успадкованих ознак буде ефективний вибір чудових індивідуальних рослин, що оцінюються в одному місці, в той час як для ознак з низьким успадкуванням відбір повинен бути оснований на середніх величинах, одержаних з повторних оцінок сімейств родинних рослин. Популярні способи селекції звичайно включають виведення чистих ліній, модифіковане виведення чистих ліній, масовий відбір і рекурентний відбір. Складний характер успадкування впливає на вибір способу селекції. Зворотне схрещування використовують для перенесення одного або декількох сприятливих генів для високо успадкованої ознаки в бажаний сорт, що культивується. Цей підхід широко використовується для селекції стійких до хвороб сортів, що культивуються. Для поліпшення кількісно 1 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 успадкованих ознак, що контролюються рядом генів, використовують різні методики рекурентного відбору. Застосування рекурентного відбору у самозапилюваних сільськогосподарських культур залежить від легкості запилення, частоти виникнення успішних гібридів від кожного запилення і числа гібридних нащадків від кожного успішного схрещування. Кожна програма селекції повинна включати періодичну об'єктивну оцінку ефективності процедури селекції. Критерії оцінки варіюють в залежності від мети і задач, але повинні включати віддачу від відбору за рік, виходячи з порівняння з відповідним стандартом, загальну цінність вдосконалених ліній, що виводяться, і ряд успішних сортів, що культивуються, одержаних на одиницю на вході (наприклад, в рік, на одиницю затрачених грошових коштів і т.д.). Перспективні вдосконалені лінії, що виводяться, ретельно тестують і порівнюють з відповідними стандартами в умовах навколишнього середовища, відповідних комерційній цільовій площі (площам) протягом трьох або більше років. Переважні лінії є кандидатами для нових комерційних сортів, що культивуються; ті, у яких відсутні невелика кількість ознак, можна використати як батьківські рослини для одержання нових популяцій для додаткового відбору. Ці процеси, які приводять до кінцевої стадії продажу і поширення, звичайно займають від восьми до дванадцяти років від початку проведення першого схрещування. Тому виведення нових сортів, що культивуються, являє собою тривалий процес, що вимагає точного довгострокового планування, ефективного використання ресурсів і мінімальної зміни напряму. Найбільш важкою задачею є ідентифікація індивідуальних рослин, які володіють перевагою на генетичному рівні, оскільки для більшості ознак дійсно цінний генотип замаскований іншими сполученими ознаками рослини або чинниками навколишнього середовища. Одним способом ідентифікації перевершуючої рослини є спостереження за її характеристиками відносно інших експериментальних рослин і відносно широко поширеного стандартного сорту, що культивується. Якщо неможливо зробити висновки по одному спостереженню, то повторні спостереження забезпечують кращу оцінку її генетичної цінності. Метою селекції рослин є створення нових, унікальних і поліпшених сортів і гібридів соняшника, що культивуються,. Селекціонер спочатку відбирає і схрещує дві або декілька батьківських ліній, з подальшими повторними самозапиленням і відбором, одержуючи безліч нових генетичних комбінацій. Теоретично, селекціонер може генерувати мільярди різних генетичних комбінацій за допомогою схрещування, самозапилення і мутацій. Селекціонер не має прямого контролю на клітинному рівні. Тому два селекціонери ніколи не виведуть однакову лінію, або навіть дуже схожі лінії, що мають однакові ознаки соняшника. Кожний рік селекціонер рослин відбирає зародкову плазму для вдосконалення наступного покоління. Цю зародкову плазму вирощують при унікальних і відмінних географічних, кліматичних і грунтових умовах, а потім проводиться додатковий відбір протягом і в кінці вегетаційного періоду. Культивовані сорти, що виводяться, є непередбачуваними. Непередбачуваність є наслідком відбору селекціонером, який відбувається в унікальному природному оточенні без контролю на рівні ДНК (з використанням традиційних процедур схрещування) і з мільйонами різних можливих генетичних комбінацій, що генеруються. Звичайний селекціонер при створенні ліній не може передбачити одержані кінцеві лінії, крім як в дуже приблизній і загальній формі. Один і той же селекціонер не може одержати один і той же культивований сорт двічі, використовуючи точно такі ж початкові батьківські рослини і однакові методики відбору. Ця непередбачуваність приводить до витрачання великої кількості коштів на дослідження для виведення нових кращих сортів соняшника, що культивуються. Виведення нових сортів соняшника, що культивуються, вимагає виведення і відбору сортів соняшника, схрещування цих сортів, і відбору кращих гібридів при схрещуванні. Гібридне насіння одержують ручним схрещуванням відібраних батьківських рослин з фертильними чоловічими рослинами або використовуючи системи чоловічої стерильності. Ці гібриди відбирають по деяких одногенних ознаках, таких як колір стручків, колір квіток, колір опушення або резистентність до гербіцидів, які вказують на те, що насіння дійсно є гібридним. Додаткові дані по батьківських лініях, а також фенотип гібрида, впливають на рішення селекціонера про те чи варто продовжувати роботу з конкретним гібридом. Для виведення сортів, що культивуються, з популяцій, що схрещуються, використовують способи селекції - виведення чистих ліній і рекурентний відбір. Програми схрещування об'єднують бажані ознаки з двох або декількох сортів, що культивуються, або різних розрізнених джерел в селекційні пули, з яких виводять культивовані сорти самозпиленням і відбором бажаних фенотипів. Нові сорти, що культивуються, оцінюють для визначення їх комерційного потенціалу. 2 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 Виведення чистих ліній звичайно використовують для поліпшення самозапилюваних сільськогосподарських культур. Дві батьківських лінії, які володіють сприятливими взаємнодоповнюючими ознаками схрещують, одержуючи F 1. Популяцію F2 одержують самозапиленням одного або декількох F1. Відбір кращих індивідуумів можна починати на популяції F2; потім, починаючи з F3, відбирають кращих індивідуумів в кращих сімействах. Для поліпшення ефективності відбору по ознаках з низьким успадкуванням повторне тестування сімейств можна починати з покоління F4. На пізніх стадіях інбридінга (а саме, F6 і F7) кращі лінії або суміші фенотипічно схожих ліній тестують на можливе застосування як нові сорти, що культивуються. Масовий і рекурентний відбір можна використати для поліпшення популяцій або само-, або перехреснозапилюваних сільськогосподарських культур. Генетично варіабельна популяція гетерозиготних індивідуумів або ідентифікується, або створюється перехресним схрещуванням декількох різних батьків. Кращі рослини відбирають, виходячи з індивідуальної переваги, видатного потомства або прекрасної комбінаційної здатності. Проводять перехресне схрещування відібраних рослин для одержання нової популяції, з якою продовжують додаткові цикли відбору. Зворотне схрещування використовують для перенесення генів для просто і високо успадкованої ознаки в бажаний гомозиготний сорт, що культивується, або інбредну лінію, які є рекурентними батьками. Джерело ознаки, яке необхідно перенести, називається донорним батьком. Очікується, що одержана рослина буде мати характеристики рекурентного батька (наприклад, сорту, що культивується) і бажану ознаку, перенесену з донорного батька. Після первинного схрещування відбирають індивідуумів, що володіють фенотипом донорного батька, і повторно схрещують (зворотне схрещування) з рекурентним батьком. Очікується, що одержана рослина буде мати характеристики рекурентного батька (наприклад, сорту, що культивується ) і бажану ознаку, перенесену з донорного батька. Метод покоління однієї насіннини в суворому значенні стосується висівання популяції, що розщеплюється, збору зразків по одній насіннині на рослину і використання цього зразка для висівання наступного покоління. Коли популяція просунеться від F 2 до бажаного рівня інбридінга, кожну з рослин, з яких одержані лінії, можна відстежити до різних F 2-індивідуумів. Число рослин в популяції знижується в кожному поколінні, внаслідок нездатності деякого насіння до проростання або деяких рослин до продукції щонайменше однієї насіннини. У результаті, не всі рослини F2, початково представлені в популяції, будуть представлені потомством при завершенні процесу поліпшення покоління. У багатонасінневому методі селекціонери соняшника звичайно збирають насіння з кожної рослини в популяції і обмолочують разом, одержуючи несортований матеріал. Частину матеріалу використовують для посіву наступного покоління, а частину залишають на зберігання. Метод іменується модифікованим методом покоління однієї насіннини. Багатонасінневий метод використовують для економії праці при зборі насіння. Значно швидше видаляти насіння машинним способом, ніж видаляти одну насіннину з кожної рослини вручну у випадку однонасінневого методу. Багатонасінневий метод також робить можливим висівати однакове число насіння популяції кожного покоління інбридінга. Збирається достатнє число насіння, щоб скомпенсувати вплив тих рослин, які не проростають, або не дають насіння. Опис інших способів селекції, що звичайно використовуються для різних ознак і сільськогосподарських культур, можна знайти в одному з декількох довідкових джерел (наприклад, Allard, 1960; Simmonds, 1979; Sneep et al., 1979; Fehr, 1987). Правильне тестування повинно детектувати основні недоліки і рівень переваги або поліпшення над існуючими сортами, що культивуються. У доповнення до демонстрації поліпшених характеристик повинен існувати попит на новий сорт, що культивується, який сумісний з промисловими стандартами, або який створює новий ринок. Введення нового сорту, що культивується, може внести додаткові витрати виробнику насіння, виробнику рослин, переробнику і споживачеві через спеціальну рекламу і продаж, змінену насіння і технологій комерційного виробництва, і використання нового продукту. У тестуванні, попередньому випуску нового сорту, що культивується, потрібно враховувати витрати на дослідження і виведення, а також технічну перевагу кінцевого сорту, що культивується. Для розмноження насінням сортів, що культивуються, повинне бути можливим легке і економічне виробництво насіння. Соняшник, Helianthus annuus L., є важливою і цінною польовою сільськогосподарською культурою. Тому постійною метою селекціонерів рослин є виведення стабільних високоврожайних сортів соняшника, що культивуються, які є агротехнічно стійкими. Поточною метою є максимізація кількості насіння, одержаного з площі, що використовується, і постачання харчуванням як тварин, так і людини. Для досягнення цієї мети селекціонер соняшника повинен 3 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 відбирати і виводити рослини соняшника, що мають ознаки, які дають в результаті кращі сорти, що культивуються. Передбачається, що нижченаведені приклади з даної галузі техніки і пов'язані з ними обмеження, є ілюстративними, а не виключними. Інші обмеження галузі техніки будуть очевидні фахівцям в даній галузі з прикладеного опису. ОПИС ВИНАХОДУ Наступні варіанти здійснення винаходу описані спільно з системами, інструментами і способами, які маються на увазі типовими і ілюстративними і не обмежують об'єм винаходу. У різних варіантах здійснення винаходу одна або декілька з описаних вище проблем зменшуються або усуваються, в той час як інші варіанти здійснення направлені на інші удосконалення. Винахід стосується нової рослини соняшника, що дає насіння, яке має низький вміст насичених жирів. Частково цей винахід стосується насіння соняшника, що має низький вміст насичених жирів, рослин або частин рослин соняшника, що дають насіння, яке має низький вміст насичених жирів, і способів одержання рослини соняшника, одержаної схрещуванням рослин соняшника, що дає насіння з низьким вмістом насичених жирів, самих з собою або з іншим сортом соняшника, що культивується, і створення за допомогою мутагенезу або трансформації варіантів рослин соняшника, що дають насіння з низьким вмістом насичених жирів. Аспекти винаходу стосуються нових рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти. Цей винахід частково стосується насіння соняшника, що має низький вміст насичених жирів і високий вміст лінолевої кислоти, рослин або частин рослин, соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти, і способів одержання рослини соняшника, одержаної схрещуванням рослин соняшника, що дає насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти, самих з собою або з іншим сортом соняшника, що культивується, і до створення за допомогою мутагенезу або трансформації варіантів рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти. Приклади насіння, що має низький вміст насичених жирів включають, але не обмежені цим, насіння, що має приблизно 2,8% або менше, приблизно 2,9% або менше, приблизно 3% або менше, приблизно 3,1% або менше, приблизно 3,2% або менше або приблизно 3,3% або менше загального вмісту разом взятих пальмітинової кислоти (16:0) і стеаринової кислоти (18:0). Приклади насіння, що має низький вміст насичених жирів і високий вміст лінолевої кислоти (18:2), включають, але не обмежені цим, насіння, що має приблизно 4,1% або менше, приблизно 5% або менше, приблизно 6% або менше, приблизно 7% або менше, приблизно 8% або менше, приблизно 9% або менше, приблизно 10% або менше, приблизно 11% або менше або приблизно 12% або менше загального вмісту разом взятих пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) і має приблизно 15%, приблизно 20%, приблизно 25%, приблизно 30%, приблизно 35%, приблизно 40%, приблизно 45%, приблизно 50%, приблизно 55%, приблизно 60%, приблизно 65%, приблизно 70% або приблизно 74% або більше лінолевої кислоти (18:2). Тому будь-які такі способи з використанням частин соняшника, які дають насіння, що має низький вміст насиченого жиру і, необов'язково, високий вміст лінолевої кислоти, є частиною цього винаходу (наприклад, самозапилення, зворотне схрещування, одержання гібридів, схрещування з популяціями і т.п.). Всі рослини, одержані з використанням рослин, що дають насіння, що мають як батька рослину з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, входять в об'єм цього винаходу. Переважно, рослину соняшника можна використати в схрещуванні з іншими, відмінними рослинами соняшника для одержання першого покоління (F1) гібридного насіння і рослин соняшника з поліпшеними характеристиками. В іншому аспекті даний винахід стосується змінених по одному або множині генів рослин соняшника, що дають насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти. Перенесений ген (гени) може, переважно, бути домінантним або рецесивним алелем. Перенесений ген (гени) може додавати такі ознаки, як резистентність до гербіцидів, резистентність до комах, резистентність до бактерійних, грибкових або вірусних захворювань, чоловічу фертильність, чоловічу стерильність, поліпшені харчові якості і промислову застосовність. Ген може являти собою природний ген соняшника або трансген, введений за допомогою генно-інженерних методик. В іншому аспекті даний винахід стосується регенерованих клітин для використання в культурі тканин рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, 4 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 необов'язково, високий вміст лінолевої кислоти. Культура тканин може бути здатна до регенерації рослин, що мають фізіологічні і морфологічні характеристики вищезазначеної рослини соняшника, що дає насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, і до регенерації рослин, що мають по суті такий же генотип, як вищезазначена рослина соняшника. Здатні до регенерації клітини в таких культурах тканин можуть являти собою зародки, протопласти, клітини меристеми, калюс, пилок, листя, пильовики, коріння, кінчики коріння, квітки, насіння, стручки або стеблини. Крім того, даний винахід стосується рослин соняшника, регенерованих з культур тканин за винаходом. В іншому аспекті даний винахід стосується способу введення бажаної ознаки в рослини соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, причому спосіб включає: схрещування рослини соняшника, що дає насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, з рослиною іншого сорту соняшника (який містить бажану ознаку), що культивується, для одержання потомства F1, причому бажана ознака вибрана з групи, що складається з чоловічої стерильності, резистентності до гербіцидів, резистентності до комах і резистентності до бактерійного захворювання, грибкового захворювання або вірусного захворювання; відбір одної або декількох рослин з потомства, які мають бажану ознаку, для одержання рослин потомства, що відбираються; схрещування рослин потомства, що відбираються ,з рослинами соняшника, які дають насіння, що має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, для одержання рослин потомства від зворотного схрещування; відбір рослин потомства від зворотного схрещування, які мають бажану ознаку і фізіологічні і морфологічні характеристики рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, для одержання відбірних рослин потомства при зворотному схрещуванні; і повторення цих стадій для одержання рослин першого покоління, що відбираються, або наступних поколінь потомства при зворотному схрещуванні, які містять бажану ознаку і всі фізіологічні і морфологічні характеристики рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти. У доповнення до типових аспектів і варіантів здійснення винаходу, описаних вище, при вивченні наступних описів будуть очевидні додаткові аспекти і варіанти здійснення. КОРОТКИЙ ОПИС КРЕСЛЕНЬ На фіг. 1 показане детальне картирування QTL (локусів кількісних ознак) низького вмісту стеаринової кислоти в інтервалі НА1875-НА1865 хромосоми LG17 (А: карти LG17 з новими маркерами (виділені блакитним кольором) і В: детальне картирування низького вмісту стеаринової кислоти в інтервалі НА1875-ORS565). На фіг. 2 показано вирівнювання послідовностей гена KASII-2 з двох батьківських ліній, на якому показані SNP і вставки і делеції (ідентифікаційні номери (ID): 333.1 (SEQ ID NO:38) і 333.2 (SEQ ID NO:39) представляють клони з ампліконів OND163R, а 332.4 (SEQ ID NO:40) і 332.5 (SEQ ID NO:41) з ампліконів H280R[1]/687R-1-8-1). На фіг. 3 показана колокалізація QTL низького вмісту пальмітинової кислоти (А) і гена жирних кислот KASIII-2 (В) на LG 5. СПОСОБИ ЗДІЙСНЕННЯ ВИНАХОДУ У нижченаведених описі і таблицях використовується ряд термінів. Для ясного і єдиного розуміння специфікації і формули винаходу, включаючи об'єм винаходу, що забезпечується цими термінами, приведені наступні визначення: Алель. Алель являє собою будь-яку одну або декілька альтернативних форм гена, які все належать однієї ознаки або характеристики. У диплоїдній клітині або організмі, два алелі даного гену займають відповідні локуси на парі гомологічних хромосом. Зворотне схрещування. Зворотним схрещуванням є процес, в якому селекціонер повторно схрещує гібридне потомство з одним з батьків, наприклад, гібрид першого покоління F 1 з одним з батьківських генотипів гібрида F1. Елітний соняшник. Сорт соняшника, що культивується, який був одержаний стабільним за деякими комерційно важливими агротехнічними ознаками, що включають стабільний вихід, що становить приблизно 100% або вище, відносно виходу контрольних сортів в тій же місцевості в той же час і при тих же умовах. В одному варіанті здійснення «елітний соняшник» означає сорт соняшника, що культивується, який був одержаний стабільним за деякими комерційно важливими агротехнічними ознаками, що включають стабільний вихід, що становить приблизно 110% або вище, відносно виходу контрольних сортів в тій же місцевості в той же час і при тих же умовах. В іншому варіанті здійснення «елітний соняшник» означає сорт соняшника, що культивується, який був одержаний стабільним за деякими комерційно важливими 5 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 агротехнічними ознаками, що включають стабільний вихід, що становить приблизно 115% або вище, відносно виходу контрольних сортів в тій же місцевості в той же час і при тих же умовах. Зародок. Зародком є маленька рослина, яка міститься в зрілому насінні. FAME-аналіз. FAME-аналіз (аналіз метилового ефіру жирних кислот) являє собою спосіб, який дозволяє точне кількісне визначення жирних кислот, які складають класи складних ліпідів. Імідазолінонова резистентність (Imi). Резистентність і/або толерантність, що додається одним або декількома генами, які змінюють ацетолактат-синтетазу (ALS), також відому як синтетаза ацетогідроксикислот (AHAS), що дозволяє ферменту чинити опір дії імідазолінону. Мутагенез. Мутагенез стосується мутагенезу рослини або частини рослини з використанням мутагену (наприклад, хімічного або фізичного агента, який збільшує частоту мутацій в цільовій рослині або частині рослини). Як необмежувальний приклад для одержання мутантних алелів в ендогенних генах рослин можна використати методику подвійного хімічного мутагенезу по Konzak, описану в патенті США № 6696294. Вміст олії. Його вимірюють як процент від цілого висушеного насіння, і він відрізняється для різних сортів. Його можна визначити з використанням різних аналітичних методик, таких як ЯМР, NIR (відображення в ближньому ІЧ-діапазоні) і екстракція по Сокслету. Загальне процентне співвідношення жирних кислот. Його визначають, екстрагуючи з насіння зразок олії, одержуючи метиловий ефір жирних кислот, присутніх в цьому зразку олії, і аналізуючи співвідношення різних жирних кислот в зразку за допомогою газової хроматографії. Композиція жирних кислот також може служити відмітною характеристикою сорту. Змінений (зміна) по одному гену. Змінена по одному гену рослина стосується рослин, які були виведені за допомогою методики селекції рослин, яка називається зворотним схрещуванням, або методами генної інженерії, в якому відновлені практично всі з бажаних морфологічних і фізіологічних характеристик сорту в доповнення до перенесеного в сорт одного гена за допомогою зворотного схрещування або генної інженерії. Стабілізований. Що відновно передається з одного покоління в інше покоління інбредних рослин одного сорту. Загальна кількість насичених жирів (TOTSAT). Загальний процентний вміст олії з насичених жирів в насінні, включаючи C12:0, C14:0, C16:0, C18:0, C20:0, C22:0 і C24:0. За одним конкретним варіантом здійснення винахід стосується нової рослини соняшника, що дає насіння, яке має низький вміст насичених жирів. Цей варіант здійснення стосується насіння соняшника, що має низький вміст насичених жирів, рослин або частин рослин соняшника, що дають насіння, яке має низький вміст насичених жирів, і способів одержання рослини соняшника, одержаного схрещуванням рослини соняшника, що дає насіння з низьким вмістом насичених жирів, самих з собою або з іншим сортом соняшника, що культивується, і до створення варіантів мутагенезом або трансформацією рослин соняшника, що дають насіння з низьким вмістом насичених жирів. Інші аспекти винаходу стосуються нових рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти. Цей варіант здійснення винаходу стосується насіння соняшника, що має низький вміст насичених жирів і високий вміст лінолевої кислоти, рослин або частин рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти, і способів одержання рослини соняшника, одержаної схрещуванням рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти, самих з собою або з іншим сортом соняшника, що культивується, і створення варіантів мутагенезом або трансформацією рослин соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти. Приклади насіння, що має низький вміст насичених жирів включають, але не обмежені цим, насіння, що має загальний вміст разом взятих пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно 2,8% або менше, приблизно 2,9% або менше, приблизно 3% або менше, приблизно 3,1% або менше, приблизно 3,2% або менше або приблизно 3,3% або менше. Приклади насіння, що має низький вміст насичених жирів і високий вміст лінолевої кислоти (18:2), включають, але не обмежені цим, насіння, що має загальний вміст разом взятих пальмітинової кислоти (16:0) і стеаринової кислоти (18:0) приблизно 6% або менше, приблизно 4,1% або менше, приблизно 5% або менше, приблизно 6% або менше, приблизно 7% або менше, приблизно 8% або менше, приблизно 9% або менше, приблизно 10% або менше, приблизно 11% або менше або приблизно 12% або менше, і маючі приблизно 15%, приблизно 20%, приблизно 25%, приблизно 30%, приблизно 35%, приблизно 40%, приблизно 45%, приблизно 50%, приблизно 55%, приблизно 60%, приблизно 65%, приблизно 70% або приблизно 74% або більше лінолевої кислоти (18:2). 6 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 Тому будь-які такі способи з використанням рослин соняшника, які дають насіння, що має низький вміст насиченого жиру і, необов'язково, високий вміст лінолевої кислоти, є частиною цього винаходу (наприклад, самозапилення, зворотне схрещування, одержання гібридів, схрещування з популяціями і т.п.). Всі рослини, одержані з використанням рослин, які дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, входять в об'єм цього винаходу. Переважно, рослину соняшника можна використати в схрещуванні з іншими, відмінними рослинами соняшника для одержання насіння і рослин гібридів соняшника першого покоління (F1) з поліпшеними характеристиками. В іншому аспекті даний винахід стосується змінених по одному або множині генів рослинам соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти. Перенесений ген (гени) може, переважно, бути домінантним або рецесивним алелем. Перенесений ген (гени) може додавати такі ознаки, як резистентність до гербіцидів, резистентність до комах, бактерійну резистентність, грибкову резистентність або резистентність до вірусних захворювань, чоловічу фертильність, чоловічу стерильність, поліпшені поживні якості і промислову застосовність. Ген може являти собою природний ген соняшника або трансген, введений за допомогою генно-інженерних методик. В іншому аспекті даний винахід стосується регенерованих клітин для використання в культурі тканин рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти. Культура тканин може бути здатна до регенерації рослин, що мають фізіологічні і морфологічні характеристики вищезазначеної рослини соняшника, що дає насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, і регенерації рослин, що мають по суті такий же генотип, як вищезазначена рослина соняшника. Здатні до регенерації клітини в таких культурах тканин можуть являти собою зародки, протопласти, клітини меристеми, калюс, пилок, листя, пильовики, коріння, кінчики коріння, квітки, насіння, стручки або стеблини. Крім того, даний винахід стосується рослин соняшника, регенерованих з культур тканин за винаходом. В іншому аспекті даний винахід стосується способу введення бажаної ознаки в рослини соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, причому спосіб включає: схрещування рослини соняшника, що дає насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, з рослиною іншого сорту соняшника, що культивується, яка містить бажану ознаку, для одержання потомства F1, причому бажана ознака вибрана з групи, що складається з чоловічої стерильності, резистентності до гербіцидів, резистентності до комах і резистентності до бактерійного захворювання, грибкового захворювання або вірусного захворювання; відбір одної або декількох рослин з потомства, які мають бажану ознаку, для одержання рослин потомства, що відбираються; схрещування рослин потомства, що відбираються, з рослинами соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, для одержання рослин потомства від зворотного схрещування; відбір рослин потомства від зворотного схрещування, які мають бажану ознаку і фізіологічні і морфологічні характеристики рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти, для одержання рослин потомства, що відбираються від зворотного схрещування; і повтор цих стадій для одержання рослин першого покоління, що відбираються, або наступних поколінь потомства від зворотного схрещування, які містять бажану ознаку і всі фізіологічні і морфологічні характеристики рослин соняшника, що дають насіння, яке має низький вміст насичених жирів і, необов'язково, високий вміст лінолевої кислоти. Придатні способи включають, але не обмежені цим, експресійні вектори, введені в тканині рослини з використанням способу прямого перенесення, такого як опосередкована мікрочастинками доставка, ін'єкції ДНК, електропорація і т.п. Експресійні вектори можна ввести в тканини рослини за допомогою опосередкованої мікрочастинками доставки з використанням пристрою Biolistic, трансформацією агробактеріями. Передбачається, що рослинитрансформанти, одержані з використанням протоплазми за винаходом, входять в об'єм цього винаходу. З приходом методик молекулярної біології, які дозволяють виділяти і характеризувати гени, що кодують певні білкові продукти, вчені в галузі біології рослин були сильно зацікавлені в зміні геному рослин відносно введення і експресії чужорідних генів, або додаткових або модифікованих версій нативних або ендогенних генів (іноді під контролем інших промоторів), з метою певної зміни ознак рослини. Такі чужорідні додаткові і/або модифіковані гени в даному описі збірно іменуються «трансгенами». За останні п'ятнадцять-двадцять років були розроблені декілька способів одержання трансгенних рослин, і даний винахід в окремих варіантах 7 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 здійснення також стосується трансформованих версій заявлених в формулі винаходу сорту або сорту, що культивується. Трансформація рослин включає конструювання експресійного вектора, який буде функціонувати в клітинах рослин. Такий вектор містить ДНК, яка включає ген під контролем регуляторного елемента або функціонально пов'язаний з ним (наприклад, промотору). Експресійний вектор може містити один або декілька таких функціонально пов'язаних комбінацій гена і регуляторного елемента. Вектор (вектори) можуть знаходитися в плазмідній формі, і їх можна використати поодинці або в комбінації з іншими плазмідами для одержання трансформованих рослин соняшника з використанням описаних нижче способів трансформації для включення трансгенів в генетичний матеріал рослини (рослин) соняшника. Експресійні вектори для трансформації соняшника: маркерні гени Експресійні вектори включають щонайменше один генетичний маркер, функціонально пов'язаний з регуляторним елементом (наприклад, промотором), який дозволяє виділити трансформовані клітини, що містять маркер, або негативною селекцією (наприклад, інгібуванням росту клітин, які не містять генів селектованого маркера), або позитивною селекцією (тобто скринінгом продукту, що кодується генетичним маркером). В галузі, пов'язаній з трансформацією, добре відома множина частовикористовуваних генів селектованих маркерів для трансформації рослин, і вони включають, наприклад, гени, що кодують ферменти, які метаболічно знешкоджують селективний хімічний агент, який може являти собою антибіотик або гербіцид, або гени, які кодують змінену мішень, нечутливу до інгібітору. У даній галузі також відомі декілька способів позитивної селекції. Одним частовикористовуваним геном селектованого маркера для трансформації рослин є ген неоміцин-фосфотрансферази II (nptII) під контролем регуляторних сигналів рослин, який додає резистентність до канаміцину. Дивіться, наприклад, Fraley et al., Proc. Natl. Acad. Sci. U.S.A., 80:4803 (1983). Іншим частовикористовуваним геном селектованого маркера є ген гігроміцин-фосфотрансферази, який додає резистентність до антибіотика гігроміцину. Дивіться, наприклад, Vanden Elzen et al., Plant Mol. Biol., 5:299 (1985). Додаткові гени селектованих маркерів бактерійного походження, які додають резистентність до антибіотиків, включають гени гентаміцин-ацетилтрансферази, стрептоміцинфосфотрансферази, аміноглікозид-3'-аденіл-трансферази і детермінанту, що визначає резистентність до блеоміцину. Дивитеся Hayford et al., Plant Physiol. 86:1216 (1988); Jones et al., Mol. Gen. Genet., 210:86 (1987); Svab et al., Plant Mol. Biol. 14:197 (1990); Hille et al., Plant Mol. Biol. 7:171 (1986). Інші гени селектованих маркерів додають резистентність до гербіцидів, таких як гліфосат, глюфозинат або бромоксиніл. Дивіться Comai et al., Nature 317:741-744 (1985); Gordon-Kamm et al., Plant Cell 2:603-618 (1990); і Stalker et al., Science 242:419-423 (1988). Інші гени селектованих маркерів для трансформації рослин мають небактерійне походження. Ці гени включають, наприклад, гени мишачої дигідрофолат-редуктази, рослинної 5-енолпірувілшикімат-3-фосфат-синтази і рослинної ацетолактат-синтази. Дивіться Eichholtz et al., Somatic Cell Mol. Genet. 13:67 (1987); Shah et al., Science 233:478 (1986); Charest et al., Plant Cell Rep. 8:643 (1990). Інший клас маркерних генів для трансформації рослин вимагає скринінга приблизно трансформованих рослинних клітин, замість прямого генетичного відбору трансформованих клітин на резистентність до токсичної речовини, такої як антибіотик. Ці гени особливо корисні для кількісної оцінки або візуалізації просторової картини експресії гена в певних тканинах і часто іменуються репортерними генами, оскільки вони можуть бути злиті з геном або регуляторною послідовністю гена для дослідження експресії гена. Частовикористовувані гени для скринінга приблизно трансформованих клітин включають гени β-глюкуронідази (GUS), βгалактозидази, люциферази і хлорамфенікол-ацетил-трансферази. Дивіться R.A. Jefferson, Plant Mol. Biol. Rep. 5:387 (1987); Teeri et al., EMBO J. 8:343 (1989); Koncz et al., Proc. Natl. Acad. Sci U.S.A. 84:131 (1987); DeBlock et al., EMBO J. 3:1681 (1984). Не так давно стали доступні способи візуалізації активності GUS in vivo, які не вимагають руйнування тканини рослин. Molecular Probes publication 2908, Imagene, T.M. Green, р. 1-4 (1993); і Naleway et al., J. Cell Biol. 115:151a (1991). Однак не було доведено, що ці способи візуалізації активності GUS in vivo корисні для одержання трансформованих клітин внаслідок своєї низької чутливості, високої фонової флуоресценції і обмежень, відповідних з використанням генів люцифераз як селектованих маркерів. Пізніше як маркер для експресії генів в прокаріотичних і еукаріотичних клітинах використали ген, що кодує зелений флуоресцентний білок (GFP). Дивіться Chalfie et al., Science 263:802 (1994). GFP і мутанти GFP можна використати як придатні для скринінга маркери. Експресійні вектори для трансформації соняшника: промотори 8 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 Включені в експресійні вектори гени повинні керуватися нуклеотидною послідовністю, що містить регуляторний елемент, наприклад, промотор. В галузі трансформації в даний момент добре відомі декілька типів промоторів, а також інші регуляторні елементи, які можна використати поодинці або в комбінації з промоторами. Термін «промотор», що використовується в даному описі, стосується області ДНК, яка розташована вище початку транскрипції (в 5'-області від початку транскрипції), і яка залучена до пізнавання і зв’язування РНК-полімерази і інших білків для ініціація транскрипції. «Рослинним промотором» є промотор, здатний ініціювати транскрипцію в рослинних клітинах. Приклади промоторів, діяльність яких залежить від стадії розвитку, включають промотори, які переважно ініціюють транскрипцію в певних тканинах, таких як листя, коріння, насіння, волокна, судини ксилеми, трахеїди або склеренхіма. Такі промотори іменуються «тканинопереважними». Промотори, які ініціюють транскрипцію тільки в певних тканинах іменуються «тканиноспецифічними». «Промотор, специфічний до типу клітин» переважно керує експресією в певних типах клітин в одному або декількох органах, наприклад, клітинах судин в корінні або листі. «Промотор, що індукується» являє собою промотор під контролем навколишнього середовища. Приклади умов навколишнього середовища, які можуть впливати на транскрипцію під контролем промоторів, що індукують ся, включають анаеробні умови або присутність світла. Тканиноспецифічні, тканинопереважні, специфічні до типу клітин і промотори, що індукують ся, складають клас «неконститутивних» промоторів. «Конститутивним» промотором є промотор, який активний при більшості умов навколишнього середовища. А. Промотори, що індукуються Промотор, що індукується, функціонально пов'язаний з геном для експресії в соняшнику. Необов'язково, промотор, що індукується, функціонально пов'язаний з нуклеотидною послідовністю, що кодує сигнальну послідовність, яка функціонально пов'язана з геном для експресії в соняшнику. З промотором, що індукується, швидкість транскрипції збільшується у відповідь на індукуючий агент. У даному винаході можна використати будь-який промотор, що індукується. Дивіться Ward et al., Plant Mol. Biol. 22:361-366 (1993). Типові промотори, що індукуються, включають, але не обмежені цим: промотори з ACEI-системи, яка відповідає на мідь (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 з маїсу, який відповідає на антидот для бензолсульфонамідних гербіцидів (Hershey et al., Mol. Gen. Genetics 227:229-237 (1991); і Gatz et al., Mol. Gen. Genetics 243:32-38 (1994)); і Tet-репрессор з Tn10 (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особливо переважним промотором, що індукується, є промотор, що відповідає на індукуючий агент, на який рослини в нормі не відповідають. Прикладом промотора, що індукується, є промотор, що індукується з гена стероїдного гормону, транскрипційну активність якого індукує глюкокортикостероїдний гормон. Schena et al., Proc. Natl. Acad. Sci. U.S.A. 88:0421 (1991). Б. Конститутивні промотори Конститутивний промотор функціонально пов'язаний з геном для експресії в соняшнику або конститутивний промотор функціонально пов'язаний з нуклеотидною послідовністю, що кодує сигнальну послідовність, яка функціонально пов'язана з геном для експресії в соняшнику. У даному винаході можна використати різні конститутивні промотори. Типові конститутивні промотори включають, але не обмежені цим: промотори з вірусів рослин, такі як промотор 35S з CaMV (Odell et al., Nature 313:810-812 (1985)); промотори актинових генів рису (McElroy et al., Plant Cell 2:163-171 (1990)); убіквітиновий промотор (Christensen et al., Plant Mol. Biol. 12:619-632 (1989), і Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)); і промотор гістона Н3 маїсу (Lepetit et al., Mol. Gen. Genetics 231:276-285 (1992), і Atanassova et al., Plant Journal 2 (3):291-300 (1992)). ALS-промотор, фрагмент XbaI/NcoI в 5'-області структурного гена ALS3 з Brassica napus (або нуклеотидна послідовність, аналогічна фрагменту XbaI/NcoI), представляє особливо придатний конститутивний промотор. Дивіться заявку РСТ WO 96/30530. В. Тканиноспецифічні або тканинопереважні промотори Тканиноспецифічний промотор функціонально пов'язаний з геном для експресії в соняшнику. Необов'язково, тканиноспецифічний промотор функціонально пов'язаний з нуклеотидною послідовністю, що кодує сигнальну послідовність, яка функціонально пов'язана з геном для експресії в соняшнику. Рослини, трансформовані представляючим інтерес геном, функціонально пов'язаним з тканиноспецифічним промотором можуть продукувати білковий продукт трансгену виключно або переважно в певній тканині. У даному винаході можна використати будь-який тканинеоспецифічний або тканинопереважний промотор. Типові тканиноспецифічні або тканинопереважні промотори включають, але не обмежені, промотором, що переважно експресується в корінні, такий як 9 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 промотор гена фазеоліну (Murai et al., Science 23:476-482 (1983) і Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985)); промотор, що специфічно експресується в листі і індукується світлом, такий як промотор генів cab або rubisco (Simpson et al., EMBO J. 4(11):27232729 (1985) і Timko et al., Nature 318:579-582 (1985)); промотор, що специфічно експресується в пильовику, такий як промотор з LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245 (1989)); промотор, що специфічно експресується в пилку, такий як промотор з Zm13 (Guerrero et al., Mol. Gen. Genetics 244:161-168 (1993)) або промотор, що переважно експресується в мікроспорах такий як промотор з apg (Twell et al., Sex. Plant Reprod. 6:217-224 (1993)). Транспорт продукованого трансгенами білка в клітинні компартменти, такі як хлоропласти, вакуолі, пероксисоми, гліоксоми, клітинна стінка або мітохондрії, або для секреції в апопласти, можна здійснити за допомогою функціонального зв’язування нуклеотидної послідовності, що кодує сигнальну послідовність з 5'- і/або 3'-областю гена, що кодує представляючий інтерес білок. У ході білкового синтезу і процесінгу направляючі послідовності на 5'- і/або 3'-кінцях структурного гена можуть визначати, в який компартмент зрештою переміститься білок, що кодується. Присутність сигнальної послідовності спрямовує поліпептид або у внутрішньоклітинну органелу, або в клітинний компартмент, або на секрецію в апопласти. У даній галузі відома множина сигнальних послідовностей. Дивіться, наприклад, Becker et al., Plant Mol. Biol. 20:49 (1992); P.S. Close, Master's Thesis, Iowa State University (1993); З. Knox et al., "Structure and Organization of Two Divergent Alpha-Amylase Genes from Barley, " Plant Mol. Biol. 9:3-17 (1987); Lerner et al., Plant Physiol. 91:124-129 (1989); Fontes et al., Plant Cell 3:483-496 (1991); Matsuoka et al., Proc. Natl Acad. Sci. 88:834 (1991); Gould et al, J. Cell. Biol. 108:1657 (1989); Creissen et al., Plant J. 2:129 (1991); Kalderon, et al., А short amino acid sequence able to specify nuclear location, Cell 39:499-509 (1984); Steifel, et al., Expression of а maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation, Plant Cell 2:785-793 (1990). Гени чужорідних білків і агротехнічні гени З використанням трансгенних рослин за даним винаходом можна одержувати чужорідний білок в промислових кількостях. Тому методики відбору і розмноження трансформованих рослин, добре вивчені в даній галузі, дають множину трансгенних рослин, які збирають стандартним чином, а потім чужорідний білок можна виділити з представляючої інтерес тканини або з всієї біомаси. Виділення білка з рослинної біомаси можна здійснити відомими способами, які обговорюються, наприклад, в Heney and Orr, Anal. Biochem. 114:92-6 (1981). В аспектах винаходу трансгенна рослина, надана для комерційного одержання чужорідного білка, є рослиною соняшника. В інших аспектах представляючою інтерес біомасою є насіння. Для відносно маленького числа трансгенних рослин, у яких спостерігається високий рівень експресії, генетичну карту можна одержати головним чином за допомогою стандартних методів аналізу RFLP (поліморфізму довжини рестрикційних фрагментів), ПЛР і SSR (повторів коротких послідовностей), які ідентифікують приблизне положення вбудованої молекули ДНК на хромосомі. Приклади методик в цьому відношенні дивитеся в Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton 269:284 (1993). Інформація про картирування відносно хромосомної локалізації корисна для захисту власності на дану трансгенну рослину. При проведенні несанкціонованого розведення і схрещування з іншою зародковою плазмою карту області вбудування можна порівняти з аналогічними картами рослин, що перевіряються, для того щоб визначити, чи мають останні спільних батьків з даною рослиною. Порівняння карт може включати гібридизацію, RFLP, ПЛР, SSR і секвенування, які є стандартним методиками. Аналогічним чином агротехнічні гени можуть експресуватися в трансформованих рослинах. Більш конкретно, за допомогою методів генної інженерії можна створити рослини для експресії різних фенотипів, що представляють агротехнічний інтерес. Приклади генів, які можна використати в цьому відношенні, включають, але не обмежені, класифіковані нижче гени. 1. Гени, які додають резистентність до шкідників або хвороб, і які кодують: А) Гени резистентності до хвороб рослин. Захист рослин часто активується специфічною взаємодією між продуктом гена резистентності до хвороби (R) в рослині і продуктом відповідного невірулентного гена (Avr) в патогені. Сорт рослини можна трансформувати клонованими генами резистентності для створення рослин, резистентних до специфічних штамів патогенів. Дивіться, наприклад, Jones et al., Science 266:789 (1994) (клонування гена Cf-9 томата, що додає резистентність до Cladosporium fulvum); Martin et al., Science 262:1432 (1993) (ген Pto томата, що додає резистентність до Pseudomonas syringae pv. tomato кодує протеїнкіназу); Mindrinos et al., Cell 78: 1089 (1994) (ген RSP2 з Arabidopsis, що додає резистентність до Pseudomonas syringae). 10 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 Б) Ген, що додає резистентність до шкідників, таких як соєва цистоутворювальна нематода. Дивіться, наприклад, заявку РСТ WO 96/30517; заявку РСТ WO 93/19181. В) Білок з Bacillus thuringiensis або його похідні, або змодельований на його основі синтетичний поліпептид. Дивіться, наприклад, статтю Geiser et al., Gene 48:109 (1986), в якій розкриті клонування і нуклеотидна послідовність гена Bt δ-ендотоксину. Більш того молекули ДНК, що кодують гени δ-ендотоксину, можна придбати в Американській колекції типових культур (АТСС), Мананасас, Віргінія, наприклад, під номерами доступу в АТСС: 40098, 67136, 31995 і 31998. Г) Лектин. Дивіться, наприклад, опис, даний в Van Damme et al., Plant Molec. Biol. 24:25 (1994), в якому розкриті нуклеотидні послідовності декількох генів манозозв’язувальних лектинів з Clivia miniata. Д) Вітамін-зв'язуючий білок, такий як авідин. Дивіться заявку РСТ US93/06487. У заявці викладається застосування авідину і авідинових гомологів в якості ларвіцидів проти комахшкідників. Е) Інгібітор ферменту, наприклад, інгібітор протеази або протеїнази або інгібітор амілази. Дивіться, наприклад, J. Biol. Chem. 262:16793 (1987) (нуклеотидна послідовність інгібітору цистеїнової протеази з рису); Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидна послідовність кДНК, що кодує протеїназний інгібітор I з тютюну); Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (нуклеотидна послідовність інгібітору альфа-амілази з Streptomyces nitrosporeus); і патент США № 5494813 (автори Hepher і Atkinson, виданий 27 лютого 1996 року). Ж) Специфічні для комах гормон або феромон, такий як екдистероїдний або ювенільний гормон, їх варіант, міметик на їх основі або їх антагоніст або агоніст. Дивіться, наприклад, опис в Hammock et al., Nature 344:458 (1990) бакуловірусної інфекції клонованої естерази ювенільного гормону, яка його інактивує. З) Специфічні для комах пептид або нейропептид, які при експресії руйнують фізіологію ураженої комахи. Наприклад, дивіться описи в Regan, J. Biol. Chem. 269:9 (1994) (експресійне клонування дозволило одержати ДНК, що кодує рецептор діуритичного гормону комах) і Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989) (ідентифікація алостатину в Diploptera puntata). Дивіться також патент США № 5266317 авторів Tomalski et al., в якому розкриті гени, що кодують специфічні для комах, паралізуючі нейротоксини. І) Специфічна для комах отрута, що в природі продукується зміями, осами і т.д. Наприклад, дивіться Pang et al., Gene 116:165 (1992), де описана гетерологічна експресія в рослинах гена, що кодує токсичний для комах пептид з скорпіону. К) Фермент, що відповідає за надмірне накопичення монотерпену, сесквітерпену, стероїду, гідроксамової кислоти, фенілпропаноїдного похідного або іншої небілкової молекули з інсектицидною дією. Л) Фермент, залучений до модифікації, в тому числі пост-трансляційної модифікації, біологічно активної молекули; наприклад, гліколітичний фермент, протеолітичний фермент, ліполітичний фермент, нуклеазу, циклазу, трансаміназу, естеразу, гідролазу, фосфатазу, кіназу, фосфорилазу, полімеразу, еластазу, хітиназу і глюканазу, або природного, або штучного походження. Дивіться заявку РСТ WO 93/02197 авторів Scott et al., в якій розкрита нуклеотидна послідовність гена калази. Молекули ДНК, які містять кодуючі хітиназу послідовності, можна одержати, наприклад, з АТСС під номерами доступу 39637 і 67152. Дивитеся також Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), в якій викладена нуклеотидна послідовність кДНК, що кодує хітиназу гусениці тютюнового бражника, і Kawalleck et al., Plant Molec. Biol. 21:673 (1993), де наведена нуклеотидна послідовність поліубіквітинового гена ubi4-2 з петрушки. М) Молекулу, яка стимулює передачу сигналу. Наприклад, дивіться розкриті в статті Botella et al., Plant Molec. Biol. 24:757 (1994), нуклеотидні послідовності для клонів кДНК кальмодуліну бобів мунг, і статтю Griess et al., Plant Physiol. 104:1467 (1994), в якій наведена нуклеотидна послідовність клону кДНК кальмодуліну маїсу. Н) Пептид з гідрофобним моментом. Дивіться заявку РСТ WO 95/16776 (в якій розкриті пептидні похідні тахіплезину, які інгібують грибкові патогени рослин) і заявку РСТ WO 95/18855 (в якій описані синтетичні антимікробні пептиди, що додають резистентність до хвороб). О) Мембранну пермеазу, яка утворює або блокує канали. Наприклад, дивіться розкриту в статті Jaynes et al., Plant Sci. 89:43 (1993) гетерологічну експресію аналога секропін-β-літичного пептиду, який наділяє трансгенні рослини тютюну резистентністю до Pseudomonas solanacearum. П) Вірус-інвазивний білок або одержаний з нього комплексний токсин. Наприклад, накопичення білків оболонки вірусу в трансформованих рослинних клітинах додає резистентність до вірусної інфекції і/або розвитку хвороби, викликаної вірусом, з якого 11 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 одержаний ген білка оболонки, а також викликаної родинними вірусами. Дивіться Beachy et al., Ann. Rev. Phytopathol. 28:451 (1990). Опосередкована білком оболонки резистентність у трансформованих рослин була одержана до вірусу мозаїки люцерни, вірусу мозаїки огірків, вірусу смугастості тютюну, вірусу Х картоплі, вірусу Y картоплі, вірусу гравіювання тютюну, вірусу погрімковості тютюну і вірусу мозаїки тютюну. Id. Р) Специфічне до комах антитіло або одержаний з нього імунотоксин. Тому антитіло, що має своєю мішенню критичну метаболічну функцію в кишечнику комах, буде інактивувати порушений фермент, вбиваючи комаху. Дивіться також Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) (ензиматична інактивація в трансгенному тютюні за допомогою продукції одноланцюжкових фрагментів антитіл). С) Специфічне до вірусу антитіло. Дивіться, наприклад, статтю Tavladoraki et al., Nature 366:469 (1993), в якій показано, що трансгенні рослини, які експресують гени рекомбінантних антитіл, захищені від атаки вірусу. Т) Зупиняючий розвиток білок, що продукується в природі патогеном або паразитом. Таким чином, грибкові ендо-α-1,4-D-полігалактуронази сприяють колонізації і вивільненню поживних речовин з рослини внаслідок солюбілізації клітинної стінки гомо-α-1,4-D-галактуроназою. Дивіться Lamb et al., Bio/Technology 10:1436 (1992). Клонування і характеристика гена, який кодує інгібуючий ендополігалактуроназу білок з квасолі, описані в статті Toubart et al., Plant J. 2:367 (1992). У) Зупиняючий розвиток білок, що продукується в природі рослинами. Наприклад, в статті Logemann et al., Bio/Technology 10:305 (1992) було показано, що трансгенні рослини, які експресують інактивуючий рибосому ген ячменю, володіли підвищеною резистентністю до грибкових захворювань. 2. Гени, які додають резистентність до гербіцидів: А) Гербіцид, який інгібує точку росту або меристему, такий як імідазолінон або сульфонілсечовина. Типові гени в цій категорії кодують мутантні ферменти ALS і AHAS, описані, наприклад, в Lee et al., EMBO J. 7:1241 (1988) і Miki et al., Theor. Appl Genet. 80:449 (1990), відповідно. Б) Гербіцид, який інгібує фотосинтез, такий як триазин (гени psbA і gs+) або бензонітрил (ген нітрилази). У статті Przibila et al., Plant Cell 3:169 (1991) описана трансформація Chlamydomonas плазмідами, що кодують мутантний ген psbA. Нуклеотидні послідовності для генів нітрилаз розкриті в патенті США № 4810648 автора Stalker, а молекули ДНК, що містять ці гени, доступні під номерами доступу АТСС - 53435, 67441 і 67442. Клонування і експресія ДНК, що кодує глутатіон-S-трансферазу, описані в Hayes et al., Biochem. J. 285:173 (1992). 3. Гени, які додають позитивну ознаку або вносять внесок в позитивну ознаку, таку як: А) Модифікований метаболізм жирних кислот, наприклад, внаслідок трансформації рослини антисмисловим геном стеарил-АСР-десатурази для збільшення вмісту стеаринової кислоти в рослині. Дивіться Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992). Б) Зниження вмісту фітату: 1) Введення кодуючого фітазу гена буде посилювати деградацію фітату, додаючи більше вільного фосфату трансформованій рослині. Наприклад, дивіться Van Hartingsveldt et al., Gene 127:87 (1993), де розкрита нуклеотидна послідовність гена фітази з Aspergillus niger. 2) Можна ввести ген, який знижує вміст фітату. Наприклад, в маїсі це можна здійснити клонуванням і потім повторним введенням ДНК, асоційованої з одним алелем, який відповідає за мутантні рослини маїсу, відмінні низьким вмістом фітової кислоти. Дивіться Raboy et al., Maydica 35:383 (1990). В) Модифікована композиція вуглеводів, що одержується, наприклад, трансформацією рослин геном, що кодує фермент, який змінює структуру гілкування крохмаля. Дивитеся Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидна послідовність гена фруктозилтрансферази з Streptococcus mutants); Steinmetz et al., Mol. Gen. Genet. 20:220 (1985) (нуклеотидна послідовність гена левансукрази з Bacillus subtilis); Pen et al., Bio/Technology 10:292 (1992) (одержання трансгенних рослин, які експрессируют α-амілазу з Bacillus lichenifonnis); Elliot et al., Plant Molec. Biol. 21:515 (1993) (нуклеотидна послідовність генів інвертаз томата); Sogaard et al., J. Biol. Chem. 268:22480 (1993) (сайт-спрямований мутагенез гена α-амілази ячменю); і Fisher et al., Plant Physiol. 102:1045 (1993) (фермент гілкування крохмалю II з ендосперми маїсу). Способи трансформації соняшника Для трансформації рослин були розроблені множина способів, включаючи протоколи біологічної і фізичної трансформації рослин. Дивіться, наприклад, Miki et al., "Procedures for Introducing Foreign DNA into Plants" в Methods in Plant Molecular Biology and Biotechnology, B.R. Glick and J.E. Thompson, Eds. (CRC Press, Inc., Boca Raton, 1993), сторінки 67-88. У доповнення 12 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 доступні експресійні вектори і способи культивування in vitro для трансформації рослинних клітин або тканин і регенерації рослин. Дивіться, наприклад, Gruber et al., "Vectors for Plant Transformation" в Methods in Plant Molecular Biology and Biotechnology, B.R. Glick and J.E. Thompson, Eds. (CRC Press, Inc., Boca Raton, 1993), сторінки 89-119. А) Опосередкована агробактеріями трансформація. Один спосіб введення експресійного вектора в рослини оснований на природній системі трансформації агробактерій. Дивіться, наприклад, Horsch et al., Science 227:1229 (1985). А. tumefaciens і А. rhizogenes є патогенними для рослин грунтовими бактеріями, які генетично трансформують рослинні клітини. Плазміди Ti і Ri з А. tumefaciens і А. rhizogenes, відповідно, несуть гени, що відповідають за генетичну трансформацію рослини. Дивіться, наприклад, C.I. Kado, Crit. Rev. Plant Sci. 10:1 (1991). Опис векторної системи на основі агробактерій і способів опосередкованого агробактеріями перенесення генів наведені в Gruber et al., supra, Miki et al., supra, і Moloney et al., Plant Cell Reports 8:238 (1989). Дивіться також патент США № 5563055 (авторів Townsend і Thomas), виданий 8 жовтня 1996 року. Б) Пряме перенесення генів. Як альтернатива для опосередкованої агробактеріями трансформації був розроблений ряд способів трансформації рослин, в сукупності генів, що іменуються прямим перенесенням. Загалом застосовним способом трансформації рослин є трансформація, опосередкована бомбардуванням мікрочастинками, в якій ДНК несуть на своїй поверхні мікрочастинки з розміром від 1 до 4 мкм. Експресійний вектор вводять в тканини рослини за допомогою біолістичного пристрою, який додає мікрочастинкам прискорення до 300600 м/с, що достатньо для проникнення через клітинну стінку і мембрани. Sanford et al., Part. Sci. Technol. 5:27 (1987); J.C. Sanford, Trends Biotech. 6:299 (1988); Klein et al., Bio/Technology 6:559-563 (1988); J.C. Sanford, Physiol. Plant 7:206 (1990); Klein et al., Biotechnology 10:268 (1992). Дивіться також патент США № 5015580 (авторів Christou, et al.), виданий 14 травня 1991 року; патент США № 5322783 (авторів Tomes, et al.), виданий 21 червня 1994 року. Іншим способом фізичної доставки ДНК в рослини є озвучення цільових клітин. Дивитеся Zhang et al., Bio/Technology 9:996 (1991). Як альтернатива, для введення експресвйних векторів в рослини використовують злиття ліпосом і сферопластів. Дивитеся Deshayes et al., EMBO J, 4:2731 (1985); Christou et al., Proc Natl. Acad. Sci. U.S.A. 84:3962 (1987). Також було опубліковане пряме поглинання ДНК протопластами з використанням CaCl2-преципітації, полівінілового спирту або полі-L-орнітину. Hain et al., Mol. Gen. Genet. 199:161 (1985), і Draper et al., Plant Cell Physiol. 23:451 (1982). Також була описана електропорація протопластів і цілих клітин і тканин. Дивитеся Donn et al., в рефератах VIIth International Congress on Plant Cell and Tissue Culture IAPTC, A2-38, стор. 53 (1990); D'Halluin et al., Plant Cell 4:1495-1505 (1992), і Spencer et al., Plant Mol. Biol. 24:51-61 (1994). Після трансформації цільових тканин соняшника експресія описаних вище генів селектованих маркерів дозволяє здійснити переважний відбір трансформованих клітин, тканин і/або рослин з використанням способів регенерації і відбору, добре відомих в даній галузі. Описані вище способи трансформації звичайно будуть використовуватися для одержання трансгенного сорту. Потім трансгенний сорт можна схрещувати з іншим (нетрансформованим або трансформованим) сортом для одержання нового трансгенного сорту. Як альтернатива, генетична ознака, яка була введена методами генної інженерії в окремий сорт соняшника, що культивується, з використанням вищеописаних методик трансформації, можна перенести в інший сорт, що культивується з використанням традиційних методик зворотного схрещування, які добре відомі в галузі селекції рослин. Наприклад, зворотне схрещування можна використати для перенесення одержаної генно-інженерним шляхом ознаки із загальнодоступного неелітного сорту в елітний сорт або з сорту, що містить чужорідний ген в своєму геномі, в сорт або сорти, які не містять цей ген. Термін «схрещування», що використовується в даному, описі може стосуватися простого схрещування Х на Y або процесу зворотного схрещування в залежності від контексту. Культура тканин соняшника Подальше одержання рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, з високим вмістом лінолевої кислоти, може відбуватися в результаті самозапилення або за допомогою культивування тканин і регенерації. Культивування різних тканин соняшника і регенерація з них рослин є відомими. Наприклад, розмноження сорту соняшника, що культивується, за допомогою культури тканин описане в патенті США 6998516. Подальше відтворювання сорту можна здійснити за допомогою культури тканин і регенерації. Культивування різних тканин соєвих бобів і регенерація з них рослин добре відомі і опубліковані в множині джерел. Як приклад можна навести патент США 6998516. Тому в іншому аспекті цей винахід стосується клітин, які при рості і диференціюванні продукують рослини 13 UA 105482 C2 5 10 15 20 25 30 35 40 45 50 55 60 соняшника, що мають насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти. Термін «культура тканини», що використовується в даному описі, вказує на композицію, що містить виділені клітини одного і того ж або різних типів, або колекцію таких клітин, впорядкованих в частині рослини. Приклади типів культур тканин включають протопласти, клітини калюса, скупчення клітин рослин і рослинні клітини, які генерують культуру тканин, інтактну в рослинах або частинах рослин, таких як зародки, пилок, квітки, насіння, стручки, листя, стеблини, коріння, кінчики коріння, пильовики і т.п. Засоби одержання і підтримки культури тканин рослин добре відомі в даній галузі. Як приклад, для одержання регенерованих рослин використали культуру тканин, що містить органи. У патентах США №№ 5959185, 5973234 5977445 і 6998516 описані деякі методики. Змінені по одному гену рослини При використанні терміну «рослина соняшника» в контексті даного винаходу він також включає будь-яку зміну гена цього сорту. Термін «зміна гена у рослини», що використовується в даному описі, стосується тих рослин соняшника, які були виведені за допомогою методики селекції, яка називається зворотним схрещуванням, або за допомогою генної інженерії, в яких відновлені практичні всі бажані морфологічні і фізіологічні характеристики сорту в доповнення до одного гена, перенесеного в сорт за допомогою методики зворотного схрещування. Способи зворотного схрещування можна використати з даним винаходом для поліпшення характеристики сорту або введення характеристики в сорт. Термін «зворотне схрещування», що використовується в даному описі, стосується повторного схрещування гібридного потомства з рекурентним батьком (наприклад, зворотне схрещування 1, 2, 3, 4, 5, 6, 7, 8 або більше разів з рекурентним батьком). Батьківську рослину соняшника, яка вносить ген для бажаної характеристики, називають «нерекурентною» або «донорним батьком». Ця термінологія відображає факт, що в протоколі зворотного схрещування нерекурентного батька використовують один раз і, відповідно, до нього не повертаються. Батьківська рослина соняшника, в яку переносять ген або гени з нерекурентного батька, відома як рекурентний батько, оскільки її використовують в декількох раундах протоколу зворотного схрещування (Poehlman & Sleper, 1994; Fehr, 1987). У типовому протоколі зворотного схрещування початковий сорт, що представляє інтерес, (рекурентного батька) схрещують з другим сортом (нерекурентним батьком), який несе один представляючий інтерес ген, який необхідно перенести. Потомство, одержане від цього схрещування, потім схрещують знов з рекурентним батьком і процес повторюють до одержання рослини соняшника, де в конвертованій рослині відновлені практичні всі бажані морфологічні і фізіологічні характеристики рекурентного батька в доповнення до одного гена, перенесеного з нерекурентного батька. Відбір відповідного рекурентного батька є важливою стадією для успішної процедури зворотного схрещування. Метою протоколу зворотного схрещування є зміна або заміщення однієї ознаки або характеристики в початковому сорті. Для здійснення цього один ген рекурентного сорту модифікують або заміняють бажаним геном нерекурентного батька, в той же час зберігаючи практично всю іншу бажану генетичну і тому бажану фізіологічну і морфологічну структуру початкового сорту. Вибір конкретного нерекурентного батька буде залежати від цілей зворотного схрещування. Однією з основних цілей є додання деякої бажаної з комерційної точки зору, агротехнічно важливої ознаки в рослину. Точний протокол зворотного схрещування буде залежати від змінних характеристики або ознаки для визначення відповідного протоколу тестування. Хоча способи зворотного схрещування спрощуються, якщо переносима характеристика являє собою домінантний алель, також можна перенести рецесивний алель. У цьому випадку може бути необхідно ввести тестування потомства для визначення успішного перенесення бажаної характеристики. Були ідентифіковані множина одногенних ознак, які звичайно не відбираються при виведенні нового сорту, але які можуть бути поліпшені методиками зворотного схрещування. Одногенні ознаки можуть бути трансгенними чи ні, приклади цих ознак включають, але не обмежені, чоловічу стерильність, восковий крохмаль, резистентність до гербіцидів, резистентність до бактерійних, грибкових і вірусних хвороб, резистентність до комах, чоловічу фертильність, поліпшений поживні якості, промислове застосування, стабільну врожайність і збільшення врожайності. Ці гени звичайно успадковуються через ядро. Декілька з цих одногенних ознак описані в патентах США №№ 5959185, 5973234 і 5977445. Цей винахід також спрямований на способи одержання рослини соняшника схрещуванням першої батьківської рослини соняшника з другою батьківською рослиною соняшника, в яких перша або друга батьківська рослини соняшника являють собою рослину соняшника, що дає насіння з низьким вмістом насичених жирів, і, необов'язково, високим вмістом лінолевої 14 UA 105482 C2 5 10 15 20 25 кислоти. Крім того, обидві перша і друга батьківські рослини соняшника можуть мати своє походження від рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти. Тому будь-які такі способи з використанням рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, є частиною цього винаходу (тобто самозапилення, зворотне схрещування, одержання гібридів, схрещування з популяціями і т.п.). Всі рослини, одержані з використанням рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, як батьки входять в об'єм цього винаходу, включаючи ті, що виведені з сортів, одержаних з рослини соняшника, що дають насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти. Переважно, щоб рослину соняшника можна було використати в схрещуванні з іншими, відмінними рослинами соняшника для одержання насіння і рослин гібридів соняшника першого покоління (F1) з поліпшеними характеристиками. Сорт за винаходом можна також використати для трансформації, в якій екзогенні гени вводяться і експресуються сортом за винаходом. Мається на увазі, що генетичні варіанти, створені або за допомогою традиційних способів селекції з використанням рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, або за допомогою трансформації рослини соняшника, що дає насіння з низьким вмістом насичених жирів і, необов'язково, високим вмістом лінолевої кислоти, за допомогою будь-якого з ряду протоколів, відомих фахівцям в даній галузі, входять в об'єм цього винаходу. ПРИКЛАДИ Даний винахід додатково описаний в наступних прикладах, які наведені для ілюстрації, і не передбачається, що вони яким-небудь чином обмежують винахід. Приклад 1: Рослини соняшника, що дають насіння з низьким вмістом насичених жирів За допомогою звичайних селекційних методик була виведена зародкова плазма соняшника з незвичайно низьким вмістом насичених жирів. Вміст олій в насінні сортів соняшника, що культивуються, наведений в таблиці 1. Таблиця 1 Зразок H757B/LS10670B-B-17-323.06 H757B/LS10670B-B-17-333.11 H757B/LS10670B-B-17-323.04 H757B/LS10670B-B-17-302.08 H757B/LS10670B-B-17-318.21 HE06EE010716.001 HE06EE010834.002 HE06EE010746.002 HE06EE010700.003 HE06EE016032.005 HE06EE016037.005 HE06EE016032.002 HE06EE010717.002 HE06EE010695.001 HE06EE010816.002 HE06EE010700.001 HE06EE010814.002 HE06EE010760.004 HE06EE010741.003 HE06EE010737.003 HE06EE016050.005 HE06EE016032.004 HE06EE010763.002 HE06EE010829.002 HE06EE010738.002 HE06EE010741.004 HE06EE010824.004 HE06EE010745.003 HE06EE010816.001 С16:0 С16:1 С18:0 С18:1 С18:2 ЗАГАЛЬНИЙ ВМІСТ НАСИЧЕНИХ ЖИРІВ С16:0+С18:0 2,34 0,09 0,48 94,18 1,51 3,39 2,82 2,47 0,11 0,51 93,62 2,11 3,42 2,98 2,24 0,09 0,53 94,25 1,49 3,45 2,77 2,70 0,13 0,50 93,26 2,24 3,67 3,2 2,45 0,11 0,54 93,62 1,73 3,68 2,99 2,17 2,31 2,40 2,48 2,42 2,25 2,40 2,44 2,48 2,34 2,48 2,46 2,54 2,34 2,33 2,41 2,44 2,43 2,53 2,78 2,42 2,35 2,81 2,52 0,11 0,11 0,11 0,13 0,10 0,08 0,10 0,10 0,12 0,12 0,14 0,10 0,11 0,11 0,13 0,08 0,11 0,11 0,13 0,15 0,11 0,10 0,11 0,11 0,82 0,65 0,72 0,57 0,64 0,75 0,70 0,82 0,66 0,88 0,65 0,79 0,63 0,93 0,96 0,73 0,63 0,78 0,70 0,62 0,88 0,80 0,68 0,80 94,29 94,74 93,87 93,46 92,86 93,06 93,00 89,76 91,93 94,10 94,31 94,11 94,07 94,51 93,53 92,57 92,49 94,28 93,26 89,75 94,10 94,14 88,66 91,45 1,41 0,82 1,03 1,78 1,82 1,71 1,72 5,51 3,20 1,24 0,89 1,19 1,16 0,73 1,12 2,67 1,80 0,98 1,84 5,22 0,61 1,15 6,32 3,77 3,63 3,68 3,68 3,78 3,82 3,86 3,87 3,88 3,88 3,88 3,90 3,91 3,92 3,93 3,93 3,94 3,94 3,94 3,95 3,96 3,96 3,97 3,98 3,98 2,99 2,95 3,12 3,05 3,06 3,00 3,09 3,26 3,14 3,22 3,13 3,24 3,16 3,26 3,29 3,13 3,07 3,21 3,23 3,40 3,30 3,15 3,48 3,32 15 UA 105482 C2 5 Приклад 2: Рослини соняшника, що дають насіння з низьким вмістом насичених жирів і високим вмістом лінолевої кислоти За допомогою звичайних селекційних методик була виведена зародкова плазма соняшника з незвичайно низьким вмістом насичених жирів. Вміст олій в насінні сортів соняшника, що культивуються, наведений в таблиці 2. Таблиця 2 Зразок С16:0 С16:1 С18:0 С18:1 С18:2 ЗАГАЛЬНИЙ ВМІСТ НАСИЧЕНИХ ЖИРІВ С16:0+С18:0 4,25 0,09 1,13 37,87 55,45 5,90 5,38 4,80 0,11 0,68 39,63 53,55 6,05 5,48 4,01 0,08 1,37 38,48 54,68 6,07 5,38 5,19 0,14 0,73 35,14 57,79 6,22 5,92 4,99 0,09 1,25 17,97 74,37 6,81 6,24 H757B/LS10670B-B-173-14.01 H757B/LS10670B-B-173-02.18 H757B/LS10670B-B-173-27.12 H757B/LS10670B-B-173-16.02 H757B/LS10670B-B-173-36.22 10 Приклад 3: Рослини соняшника, що дають насіння з низьким вмістом насичених жирів За допомогою звичайних селекційних методик була виведена зародкова плазма соняшника з незвичайно низьким вмістом насичених жирів. Вміст олій в насінні сортів соняшника, що культивуються, наведений в таблиці 3. Таблиця 3 C16:0 C16:1 C18:0 C18:1 C18:2 ЗАГАЛЬНИЙ ВМІСТ НАСИЧЕНИХ ЖИРІВ NuSun/без насичених жирів NS1982.16/OND163R-1-05 NS1982.8 2,29 2,09 0,05 0,08 0,65 0,55 67,37 79,40 28,19 15,99 3,48 3,10 Без насичених жирів/високий вміст олеїнової кислоти NS1982.8-03 NS1982.8 H117R[4]//H757B/LS10670B///NS1982.6-2-023-1-12-076 1,60 1,63 1,79 0,03 0,07 0,05 0,37 0,41 0,29 95,13 94,81 95,30 1,48 1,26 0,84 2,33 2,48 2,57 5,29 3,76 0,07 0,07 0,73 0,80 18,19 34,97 74,43 58,62 6,41 5,29 3,13 0,02 2,07 36,03 56,65 6,23 Низький вміст стеаринової кислоти NS1982.8/OND163R-2-12-009 H117R[4]//H757B/LS10670B///NS1982.6-2-023-1-12-038 OID263R/NS1982.8-4-12-002 2,75 1,90 3,08 0,66 0,04 0,12 0,25 0,27 0,27 92,95 95,03 93,54 1,99 1,00 1,48 3,43 2,65 3,87 Низький вміст пальмітинової кислоти H251B[2]/IAST-4=1=100//NS1982.16-11-39-041 NS1982.14-08 NS1982.16 1,47 1,51 1,52 0,24 0,02 0,06 2,59 2,24 1,05 92,59 92,84 94,37 0,65 1,35 0,85 5,42 4,90 3,39 Дуже високий вміст олеїнової кислоти H117R[4]//H757B/LS10670B//NS1982.6-2-023-1-12-076 NS1982.8/OND163R-2-12-059 ON3351B/NS1982.8-1-04 1,79 1,87 2,04 0,05 0,10 0,03 0,29 0,44 0,50 95,30 95,22 95,20 0,84 0,97 0,70 2,57 2,76 3,08 Зразок Низький вміст насичених жирів/високий вміст лінолевої кислоти CND117R/NS1982.8-3-06 OI1601B[2]//H757B/LS10670B[1]///NS1982.6=В-3-04 CN2343B/4/CN2343B[2]//H757B/LS10670B///NS1982.11#1#13-11 15 Як видно з таблиці 3, дані показують, що олія насіння має загальний вміст насичених жирів, знижений практично до 2,33% при високому вмісті олеїнової кислоти (>80%), відсутність насичених жирів (
ДивитисяДодаткова інформація
Назва патенту англійськоюLow saturated-fat sunflower and associated methods
Автори російськоюGerdes, James, Todd, Kahl, Charles, James, Erickson, Angela, Lee, Benson, Robert, Martin
МПК / Мітки
МПК: A01H 5/10
Мітки: соняшник, вмістом, низьким, насичених, жирів
Код посилання
<a href="https://ua.patents.su/41-105482-sonyashnik-z-nizkim-vmistom-nasichenikh-zhiriv.html" target="_blank" rel="follow" title="База патентів України">Соняшник з низьким вмістом насичених жирів</a>
Продукти харчування з низьким вмістом насичених і транс-ізомерних ненасичених жирів

Номер патенту: 95101
Опубліковано: 11.07.2011
Автори: Юшіода Тошіо, Клеєневерк Бернард
МПК: A23G 3/40, A23G 1/36, A23D 7/02, A23G 1/30
Мітки: ненасичених, транс-ізомерних, низьким, продукти, вмістом, жирів, харчування, насичених
Формула / Реферат:
1. Структурований продукт харчування з безперервною жировою фазою, де продукт харчування містить по відношенню до загальної маси продукту:a) менше 30 ваг. % насичених жирних кислот,b) 20-100 ваг. % тригліцеридної композиції,c) 0-80 ваг. % наповнювача,d) менше 15 ваг. % води,в якому тригліцеридна композиція містить по відношенню до маси тригліцеридної композиції:e) менше 45 ваг. % насичених жирних...
Продукти харчування з низьким вмістом насичених і транс-ізомерних ненасичених жирів

Номер патенту: 92931
Опубліковано: 27.12.2010
Автори: Вербеєкк Сабріна, Клеєневерк Бернард, Юшіода Тошіо
МПК: C11C 3/00, A23D 7/00, A23G 1/00
Мітки: транс-ізомерних, вмістом, жирів, харчування, насичених, ненасичених, продукти, низьким
Формула / Реферат:
1. Структурований продукт харчування з безперервною жировою фазою, де продукт харчування містить по відношенню до загальної маси продукту:а) 0-35 ваг. % насичених жирних кислот,b) 20-100 ваг. % тригліцеридної композиції,с) 0-80 ваг. % наповнювача,d) 0-15 ваг. % води,в якому тригліцеридна композиція містить по відношенню до маси тригліцеридної композиції:e) менше 50 ваг. % насичених жирних...
Спосіб виробництва гранул цеоліту х з низьким вмістом діоксиду кремнію і з низьким вмістом інертного зв’язуючого

Номер патенту: 61932
Опубліковано: 15.12.2003
Автор: Пле Домінік
МПК: C01B 39/00, B01J 20/18
Мітки: гранул, інертного, вмістом, кремнію, цеоліту, низьким, діоксиду, виробництва, спосіб, зв'язуючого
Формула / Реферат:
1. Спосіб виробництва цеолітових тіл, які складаються принаймні на 95% з цеоліту Х з низьким вмістом діоксиду кремнію, який включає такі стадії:(а) агломерація порошку цеоліту Х з низьким вмістом діоксиду кремнію за допомогою зв'язуючого, що містить принаймні 80% глини, що може перетворюватись на цеоліт,(б) формування суміші, отриманій за п. (а),(в) висушування і подальша кальцинація при температурі від 500
Спосіб та плавильна установка для одержання сталі з високим вмістом марганцю і низьким вмістом вуглецю

Номер патенту: 82962
Опубліковано: 26.05.2008
Автори: Вайшедель Вальтер, Розе Лутц
МПК: C21C 5/35, C21C 5/28, C22C 33/04, C22C 38/04, C22C 35/00, C22C 38/00
Мітки: вуглецю, плавильна, установка, високим, вмістом, спосіб, сталі, одержання, низьким, марганцю
Формула / Реферат:
1. Плавильна установка для одержання сталі з високим вмістом марганцю і низьким вмістом вуглецю, яка включає конвертер (6а) з засобами подачі до нього розплавів чавуну або вуглецевої сталі та феромарганцю, шлакоутворюючих і супутніх елементів, і розміщену за конвертером (6а) по ходу технологічного процесу піч-ківш (12), яка відрізняється тим, що перед конвертером (6а) передбачена відновна піч (16) для феромарганцю, а також шахтна піч (17) для...
Жирова композиція з покращеною текстурою і ступенем кристалізації, що містить жири з низьким вмістом транс-жирних кислот

Номер патенту: 94030
Опубліковано: 11.04.2011
Автор: Клеєневерк Бернард
МПК: A23D 9/007, A23D 9/02
Мітки: кислот, транс-жирних, покращеною, жири, композиція, текстурою, кристалізації, вмістом, містить, жирова, ступенем, низьким
Формула / Реферат:
1. Жирова композиція, яка відрізняється тим, що складається з деякої кількості першого жиру, що має вміст транс-жирних кислот менше 30 ваг. % від ваги першого жиру, співвідношення насичених жирних кислот С-16/С-18 у ваговій пропорції, що складає принаймні 6, і вміст твердого жиру при 20 °С принаймні 35 ваг. %, і 0,1-5 ваг. % від загальної ваги жирової композиції прискорювача кристалізації жиру, який являє собою суміш тригліцеридів, що має...
Попередній патент: Переробка газоподібних вуглеводнів
Наступний патент: Спосіб переведення дрібнокускового мінерального карбонату на придатну для випалювання форму
Випадковий патент: Пристрій для проведення гри "магалла"