Спосіб одержання рекомбінантних штамів дріжджів saccharomyces cerevisiae з підвищеною активністю лужної фосфатази
Номер патенту: 104364
Опубліковано: 27.01.2014
Автори: Сибірний Андрій Андрійович, Дмитрук Костянтин Васильович, Семків Марта Віталіївна






Формула / Реферат
Спосіб одержання рекомбінантних штамів дріжджів Saccharomyces cerevisiae з підвищеною активністю лужної фосфатази, який відрізняється тим, що в геном дріжджів S. cerevisiae вводять вектор для мультикопійної інтеграції, який містить експресійну касету, що складається з промотору гена алкогольдегідрогенази (ADH1), відкритої рамки зчитування гена лужної фосфатази (РHО8) та термінатора гена цитохрому С (CYC1), та модифікований ген kanМХ4 (виконує роль селективного маркера), фланковані d-послідовностями.
Текст
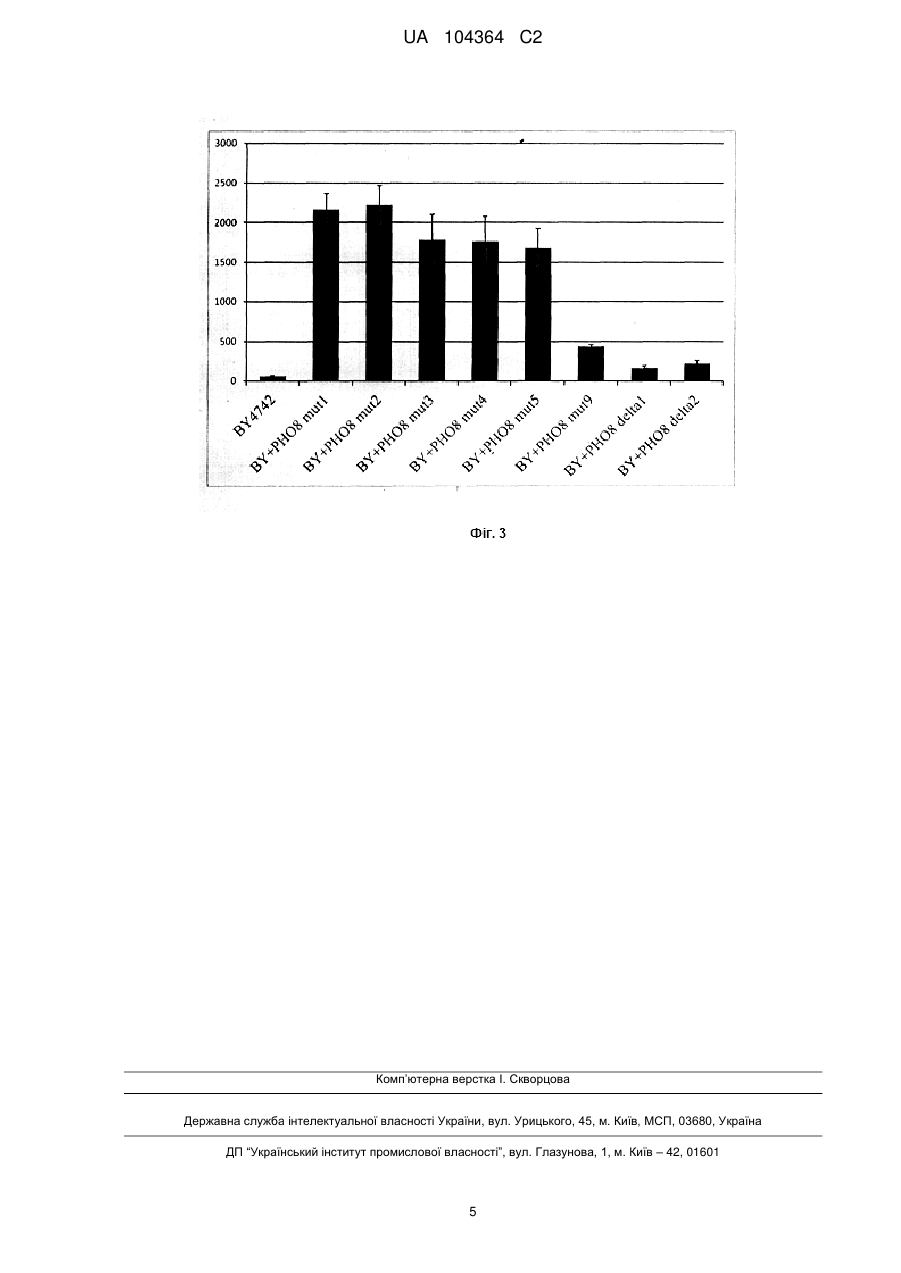
Реферат: Винахід належить до способу одержання рекомбінантних штамів дріжджів S. cerevisiae з підвищеною активністю лужної фосфатази, де підвищення активності здійснюють за допомогою введення вектора для мультикопійної інтеграції, який містить експресійну касету, що складається з промотору гена алкогольдегідрогенази (ADH1), відкритої рамки зчитування гена UA 104364 C2 (12) UA 104364 C2 лужної фосфатази РНО8, термінатора гена цитохрому С (CYC1), та селективного маркера (ген kanMX4). UA 104364 C2 5 10 15 20 25 30 35 40 45 50 55 Винахід належить до галузі біотехнології і є способом отримання рекомбінантних штамів дріжджів Saccharomyces cerevisiae з підвищеною активністю лужної фосфатази за допомогою мультикопійної інтеграції гена РНО8 в геном цих дріжджів. Лужна фосфатаза - це фермент, що здатний здійснювати гідроліз дифосфатного зв'язку у різних речовинах, зокрема в АТФ - сполуці, що необхідна для синтезу компонентів дріжджових клітин. Припускається, що зниження внутрішньоклітинного рівня АТФ у клітинах дріжджів S.cerevisiae призведе до підвищення продуктивності алкогольної ферментації за рахунок того, що менша кількість спожитої глюкози використовуватиметься для накопичення дріжджової біомаси, і більша її кількість перетворюватиметься до кінцевого продукту - етанолу [1]. Відомо, що здійснювалася експресія модифікованих форм гена РHО8 під контролем сильного конститутивного промотору з метою використання отриманих рекомбінантних штамів для, дослідження процесів автофагії у клітинах дріжджів, як наприклад описано в [2], проте модифікований ген вводився в клітини S. cerevisiae або в складі реплікативних плазмід, які хоч і забезпечують високу копійність гена в клітині, проте легко втрачаються у неселективних умовах, або в складі інтегративних векторів, які є стабільними, проте низькокопійними. Також відомо, що для забезпечення стабільної мультикопійної інтеграції гена в геном дріжджів S. cerevisiae використовуються вектори, що містять δ-послідовності, δ-послідовності це довгі кінцеві повтори нуклеотидів, що входять до складу ретротрансиозонів Ту1 і Ту2, що вбудовуються в геномну ДНК дріжджів. Копії цих нуклеотидних послідовностей залишаються у вихідній ділянці молекули ДНК після того, як відбулася транспозиція Ту-елемента в іншу ділянку, тому в геномі дріжджів присутня також велика кількість ізольованих δ-послідовностей. Загалом налічується близько 425 таких послідовностей у геномній ДНК S. cerevisiae [3]. Тому вектор, що містить δ-послідовності, забезпечуватиме інтеграцію фланкованої цими послідовностями ділянки ДНК у δ-локуси в геномній ДНК за рахунок гомологічної рекомбінації. При використанні такого вектора для трансформації можна отримати рекомбінантні штами, що містять від 1 до 8-10 копій вектора, інтегрованого у геномну ДНК. Для того, щоб отримувати лише штами з мультикопійною інтеграцією вектора, можна використовувати модифіковані селективні маркери, що здатні підтримувати ріст клітин дріжджів у селективних умовах лише в тому випадку, якщо вони присутні в геномі в кількох копіях. Найбільш близьким до запропонованого є спосіб отримання рекомбінантних штамів S. cerevisiae з високою копійністю досліджуваного гена за рахунок використання вектора, що містить нуклеотидні послідовності, гомологічні послідовностям рРНК-локусів S. cerevisiae і модифікований селективний маркер - ген Ieu2-d, що має вкорочену промоторну ділянку (близько 50 нуклеотидів) і, за рахунок цього, знижений рівень експресії [4]. Однак, використання гена Ieu2-d, як селективного маркера, потребує попереднього відбору штамів S. cerevisiae, нездатних рости на середовищі без лейцину, тобто з пошкодженим геном LEU2. Тому такий підхід не може бути застосовано для трансформації промислових штамів дріжджів S. cerevisiae, які переважно є прототрофними. В основу винаходу поставлено задачу отримати рекомбінантні штами дріжджів S. cerevisiae із підвищеною активністю лужної фосфатази за рахунок експресії гена РНО8 під контролем сильного конститутивного промотору гена алкогольдегідрогенази (ADH1) у складі вектора для мультикопійної інтеграції в геномну ДНК дріжджів. Поставлена задача вирішується тим, що у способі отримання рекомбінантних штамів дріжджів S. cerevisiae з підвищеною активністю лужної фосфатази, згідно з винаходом, в геном дріжджів S. Cerevisiae вводиться сконструйований нами вектор для мультикопійної інтеграції, який містить експресійну касету, що складається з промотору гена алкогольдегідрогенази (ADH1), відкритої рамки зчитування (ВРЗ) гена лужної фосфатази (РНО8) та термінатора гена цитохрому С (CYC1), та модифікований ген kanМХ4 (виконує роль селективного маркера), фланковані δ-послідовностями, що забезпечує підвищення ферментативної активності лужної фосфатази. У запропонованому способі використовуються методи: полімеразна ланцюгова реакція (ПЛР), конструювання рекомбінантних плазмід, виділення плазмід з Escherichia coli, рестрикційний аналіз, електрофорез в агарозному гелі, трансформація Е. coli методом електропорації, описані в [5]. Трансформацію S. cerevisiae проводять, як описано в [6]. Виділення сумарної ДНК з трансформантів S. cerevisiae проводиться як описано в [7]. Визначення питомої активності лужної фосфатази проводиться як описано в [8]. Дот-блот гібридизацію проводиться як описано в [5]. Спосіб отримання рекомбінантних штамів дріжджів S. cerevisiae з підвищеною активністю лужної фосфатази ілюструється графічними матеріалами: 1 UA 104364 C2 5 10 15 20 25 30 35 На Фіг. 1. зображено лінійну схему плазміди pUC57-deltal_2-ADHpr-PHO8-CYCt-kanMXmut, де фрагмент, що містить ген РНО8 S. cerevisiae, позначено прямокутником з колами; фрагмент, що містить промотор ADH1 S. cerevisiae, позначено прямокутником з ромбами; фрагмент, що містить термінатор CYC1 S. cerevisiae, позначено прямокутником з хрестиками; фрагмент, що містить модифікований селективний маркер kanМХ4, позначено сірим прямокутником; фрагменти, що містять δ-послідовності ретротранспозона Ту1, позначено чорними стрілками; тонкою чорною лінією позначено бактерійну частину вектора. На Фіг. 2. зображено дот-блот гібридизацію, що проводилася для визначення кількості копій гена РНO8 у геномі рекомбінантних штамів, де BY4742 - штам дикого типу, що має 1 копію гена РНО8 в геномній ДНК; BY+PHO8 mut - рекомбінантні штами, що містять вектор pUC57-deltal_2-ADHpr-PHO8-CYCtkanMXmut з модифікованим геном kanМХ4\ BY+PHO8 delta - рекомбінантні штами, що містять вектор з немодифікованим геном kanМХ4. На Фіг. 3. зображено питому активність лужної фосфатази, виражену в мкмоль продукту/мг білка*хв., в безклітинних екстрактах рекомбінантних штамів S. cerevisiae в порівнянні зі штамом дикого типу. Позначення штамів такі ж, як на Фіг. 2. Спосіб отримання рекомбінантних штамів дріжджів S. cerevisiae з підвищеною активністю лужної фосфатази здійснюють кількома етапами: Етап 1. Конструювання плазміди, що містить δ-послідовності та ген РНO8 під промотором ADH1 На початковому етапі геномну ДНК S. cerevisiae BY4742 використовують як матрицю для ампліфікації δ-послідовностей за допомогою ПЛР: частину послідовності YJRWdeltal2 [9] розміром 154 пн ампліфікують з допомогою праймерів SM16/SM17 (Таблиця), іншу частину розміром 180 пн ампліфікують з допомогою праймерів SM18/SM19. В подальшому обидва фрагменти об'єднують з допомогою ПЛР з використанням праймерів SM16/SM19, обробляють рестриктазами FxoRI та Hindlll і клопують у плазмідний вектор pUC57. Сконструйовану плазміду позначено pUC57-deltal_2. Фрагмент ДНК розміром 807 пн, що відповідає промотору гена ADH1 (кодує алкогольдегідрогеназу), ампліфікують з геномної ДНК штаму S. cerevisiae BY4742 за допомогою праймерів Ко419 і Ко420. Фрагмент ДНК розміром 269 пн, що відповідає термінатору гена CYC1 (що кодує цитохром С), ампліфікують з геномної ДНК штаму S. cerevisiae BY4742 за допомогою праймерів Ко453 та Ко454. Потім ці два фрагменти об'єднують з допомогою ПЛР з використанням праймерів Ко419 і Ко454. Отриманий фрагмент ДНК обробляють ендопуклеазами рестрикції Sail і Xmal і клонують по відповідних сайтах у плазміду pUC57deltal_2. Отриману плазміду позначено pUC57-deltal_2-ADHpr-CYCt. Фрагмент ДНК розміром 1701 пн, що відповідає ВРЗ гена РHO8 S. cerevisiae, ампліфікують з використанням праймерів Ко508 і Ко509 і геномної ДНК BY4742 як матриці. Після очистки цей фрагмент обробляють ендонуклеазами рестрикції ВаmHІ і NotI і клонують в плазміду pUC57-deltal_2-ADHpr-CYCl. Отриманий вектор названо pUC57-deltal_2-ADHpr-PHO8-CYCt. Таблиця Перелік праймерів, які використовують на 1 та 2 етапах способу 40 Етап 2. Модифікацій гена kanМХ4, що забезпечує резистентність до антибіотика і енстицину. 2 UA 104364 C2 5 10 15 20 25 30 35 40 45 50 55 60 Ген kanМХ4 застосовується як гетерологічний домінантний селективний маркер для S.cerevisiae. Наявність щонайменше однієї копи цього гена в геномній ДНК дріжджів забезпечує їх резистентність до антибіотика генетицину (в концентрації 200 мг/л для S. cerevisiae). Було поставлено завдання модифікувати ген kanМХ4 таким чином, щоб він працював менш ефективно. Тоді для селекції дріжджових трансформантів необхідна наявність багатьох копій модифікованого селективного маркера в геномі відповідних штамів. Селекцію модифікованого гена kanМХ4 здійснюють в клітинах Е. соlі. Для цього спочатку визначають мінімальну концентрацію генетицину, що здатна інгібувати ріст культури Е. соlі - це 2 мг генетицину на 1 л середовища. В подальшому проводять ПЛР з використанням праймерів Ко446 та Ко447 (див. табл.) на матриці плазміди pRS303K, що містить у своєму складі ген kanМХ4, за умов, що сприяють виникненню великої кількості помилок при копіюванні ланцюга ДНК (висока 2+ 2+ концентрація іонів Mg , наявність іонів Мn , непропорційні концентрації нуклеотидтрифосфатів у реакційній суміші, використання ДНК-полімерази з низькою точністю зчитування). Сукупність ампліфікованих фрагментів ДНК, що містять у своєму складі різноманітні нулеотидні заміни, обробляють ендонуклеазою рестрикції ХbаІ і клонують у відповідний сайт плазміди pUC57. Відбір трансформантів Е. соlі, що містять плазміду зі вставкою, проводять на середовищі з ампіциліном (100 мг/л) та генетицином (2 мг/л). Отримані колонії трансформантів за допомогою методу реплік переносять на середовище з вищою концентрацією генетицину (10 мг/л) і відбирають трансформанти, що не здатні рости при такій концентрації антибіотика. У подальшому з одного з відібраних трансформантів Е. соlі отримують плазміду, що містить у своєму складі модифікований ген kanМХ4. Модифікований ген kanМХ4 у складі SacI/SmaIфрагмента, обробленого Т4-ДНК-полімеразою для утворення "тупих" кінців клонують у Хbалінеаризований та оброблений Т4-ДНК-полімеразою вектор pUC57-deltal_2-ADHpr-PHO8-CYCt. Отриману плазміду позначено pUC57-deltal_2-ADHpr-PHO8-CYCt-kanMXmut (Фіг. 1). Також для порівняння конструюють вектор, що містить нативний ген kanМХ4, його позначено pUC57deltal_2-ADHpr-PHO8-CYCt-kanMX. Етап 3. Конструювання рекомбінантних штамів S. cerevisiae. Отримані на попередньому етапі 'роботи вектори pUC57-deltal_2-ADHpr-PHO8-CYCtkanMXmut та pUC57-deltal_2-ADНpr-PHO8-CYCt-kanMX обробляють ендонуклеазою рестрикції Ahdl (при цьому видаляється частина вектора, що відповідає плазміді pUC57) і використовують для трансформації штаму S. cerevisiae BY4742. Селекцію генетицин-резистентних трансформантів проводять на середовищі YPD з додаванням 2G0 мг/л генетицину. Наявність в геномі трансформантів експресійної касети ADHpr-PHO8 перевіряють за допомогою ГІЛР з використанням відповідної пари праймерів (Ко419 та Ко509) (див. табл.). Визначення копійності гена РНО8 в геномі рекомбінантних штамів здійснюється за допомогою дот-блот гібридизації (Фіг. 2). Вектор pUC57-deltal_2-ADНpr-PНO8-CYCt-kanMX забезпечує високу частоту 3 трансформації S. cerevisiae, ~10 трансформантів/мкг ДНК, проте включається в геном найчастіше в 1-2 копіях, а вектор pUC57-deltal_2-ADHpr-PНO8-CYCt-kanMXmul, що забезпечує 1 значно нижчу частоту трансформації, ~ 10 трансформантів/мкг ДНК, дозволяє отримати трансформанти, що містять від 3 до 7-9 додаткових копій гена РНО8. Визначення питомої активності лужної фосфатази у рекомбінантних штамів (Фіг. 3) показало, що штами, які містять вектор pUC57-deltal_2-ADHpr-PHO8-CYCt-kanMXmut мають від 7 до 35 разів вищу відповідну ферментативну активність у порівнянні з реципієнтним штамом BY4742 та у 2-10 разів вищу у порівнянні зі штамами, що містять вектор pUC57-deltal_2-ADHpr-PHO8-CYCt-kanMX. Значення активності лужної фосфатази в отриманих рекомбінантних штамів S. cerevisiae добре корелюють з кількістю додаткових копій гена РНО8 у їх геномі. Отже, вектор pUC57-deltal_2ADHpr-PHO8-CYCt-kanMXmut забезпечує мультикопійну інтеграцію гена РНО8 у геном S. cerevisiae. Джерела інформації:; 1. Stefan de Kok et al. // FEMS Yeast Research-2012. - Vol. 12. - P. 387-397. 2. Corey L. etal.// Journal of Cell Science-1998-vol.111-P. 2455-2464. 3. Lee F.W.F. et al. // Appl Microbiol Biotechnol-1997-vol.48-P. 339-345. 4. Romanos М.Л. et al.// Yeasl-1992-vol.8-P. 423-488. 5. Sambrook J., Fritsh E. F., Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. 1989. 6. Gietz RD et al. // Methods in Molecular Biology-2006-vol.313-P.107-120. 7. Wach A., Pick H., Philipsen P. In: Molecular Genetics of Yeast. A Practical Approach (Johnston, J.R.J Ed.), IRL Press, Oxford.-1994. - P. 1-16. 8. KanekoY. etal.// Мої. Cell Biol.-1982-vol.2-P. 127-137. 3 UA 104364 C2 9. Saccharomyces Genome Database//www.yeastgenome.org/ 5 10 ФОРМУЛА ВИНАХОДУ Спосіб одержання рекомбінантних штамів дріжджів Saccharomyces cerevisiae з підвищеною активністю лужної фосфатази, який відрізняється тим, що в геном дріжджів S. cerevisiae вводять вектор для мультикопійної інтеграції, який містить експресійну касету, що складається з промотору гена алкогольдегідрогенази (ADH1), відкритої рамки зчитування гена лужної фосфатази (РHО8) та термінатора гена цитохрому С (CYC1), та модифікований ген kanМХ4 (виконує роль селективного маркера), фланковані -послідовностями. 4 UA 104364 C2 Комп’ютерна верстка І. Скворцова Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 5
ДивитисяДодаткова інформація
Автори англійськоюDmytruk Kostiantyn Vasyliovych, Sybirnyi Andrii Andriiovych
Автори російськоюДмитрук Константин Васильевич, Сибирный Андрей Андреевич
МПК / Мітки
МПК: C12N 15/69
Мітки: підвищеною, активністю, cerevisiae, дріжджів, штамів, одержання, фосфатази, saccharomyces, лужної, спосіб, рекомбінантних
Код посилання
<a href="https://ua.patents.su/7-104364-sposib-oderzhannya-rekombinantnikh-shtamiv-drizhdzhiv-saccharomyces-cerevisiae-z-pidvishhenoyu-aktivnistyu-luzhno-fosfatazi.html" target="_blank" rel="follow" title="База патентів України">Спосіб одержання рекомбінантних штамів дріжджів saccharomyces cerevisiae з підвищеною активністю лужної фосфатази</a>
Спосіб отримання рекомбінантних штамів дріжджів saccharomyces cerevisiae з підвищеною активністю лужної фосфатази

Номер патенту: 79020
Опубліковано: 10.04.2013
Автори: Дмитрук Костянтин Васильович, Семків Марта Віталіївна, Сибірний Андрій Андрійович
МПК: C12N 15/69, C12N 15/81
Мітки: saccharomyces, активністю, штамів, фосфатази, спосіб, лужної, cerevisiae, дріжджів, отримання, підвищеною, рекомбінантних
Формула / Реферат:
Спосіб отримання рекомбінантних штамів дріжджів Sacchromyces cerevisiae з підвищеною активністю лужної фосфатази, який відрізняється тим, що в геном дріжджів S. cerevisiae вводять вектор для мультикопійної інтеграції, який містить експресійну касету, що складається з промотору гена алкогольдегідрогенази (ADН1), відкритої рамки зчитування гена лужної фосфатази (РНО8) та термінатора ієна цитохрому С (CYC1), та модифікований ген kаnМХ4 (виконує...
Штам дріжджів saccharomyces cerevisiae для одержання сушених хлібопекарських дріжджів

Номер патенту: 36624
Опубліковано: 16.06.2003
Автори: Шустікова Наталія Євгенівна, Левандовський Леонід Вікторович, Рудніченко Людмила Вікторівна, Олійнічук Сергій Тимофійович, Коваль Катерина Олександрівна, Хоменко Анатолій Іванович, Ткаченко Алла Феодосіївна
МПК: C12R 1/865, C12N 1/18
Мітки: дріжджів, cerevisiae, saccharomyces, хлібопекарських, одержання, штам, сушених
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ІMB У-5009 для одержання сушених хлібопекарських дріжджів.
Штам дріжджів saccharomyces cerevisiae хл-1 для одержання хлібопекарських дріжджів

Номер патенту: 30677
Опубліковано: 15.03.2002
Автори: Рудніченко Людмила Вікторівна, Олійнічук Сергій Тимофійович, Шустікова Наталія Євгенівна, Коваль Катерина Олександрівна, Левандовський Леонід Вікторович, Ткаченко Алла Феодосіївна
МПК: C12R 1/865, C12N 1/18
Мітки: дріжджів, хл-1, saccharomyces, cerevisiae, штам, хлібопекарських, одержання
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ХЛ-1 для одержання хлібопекарських дріжджів.
Штам дріжджів saccharomyces cerevisiae imb-у-5016 для одержання пресованих хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів

Номер патенту: 47821
Опубліковано: 15.01.2004
Автори: Рудніченко Людмила Вікторівна, Бейко Наталія Євгенівна, Хоменко Анатолій Іванович, Ткаченко Алла Феодосіївна, Левандовський Леонід Вікторович, Коваль Катерина Олександрівна
МПК: C12R 1/865, C12N 1/18
Мітки: спирту, штам, дріжджів, хлібопекарських, одержання, saccharomyces, imb-у-5016, cerevisiae, виробництві, двопродуктовому, пресованих
Формула / Реферат:
Штам дріжджів Saccharomyces cerevisiae ІМВ-У-5016 для одержання хлібопекарських дріжджів в двопродуктовому виробництві спирту і хлібопекарських дріжджів.
Спосіб одержання поверхневого антигену віруса гепатиту в за допомогою рекомбінантних штамів дріжджів hansenula polymorpha з пошкодженою катаболітною репресією

Номер патенту: 73449
Опубліковано: 25.09.2012
Автори: Красовська Олена Сильвестрівна, Стасик Олег Володимирович, Сибірний Андрій Андрійович
МПК: C12P 7/06
Мітки: пошкодженою, репресією, допомогою, поверхневого, polymorpha, дріжджів, одержання, гепатиту, рекомбінантних, спосіб, катаболітною, антигену, hansenula, віруса, штамів
Формула / Реферат:
Спосіб одержаного поверхневого антигену вірусу гепатиту В за допомогою рекомбінантних штамів дріжджів Hansenula Polymorpha з пошкодженою катаболітною репресією, який відрізняється тим, що експресія поверхневого антигена вірусу гепатиту В здійснюється мутантними штамами дріжджів Н. polymorpha з пошкодженою катаболітною репресією, здатними синтезувати поверхневий антиген на середовищі з глюкозою або ксилозою, а селекція мультикопійних...
Попередній патент: Спосіб активації ендотелію ізольованого фрагмента судини у моделюванні ендотелійзалежних процесів та пристрій для їх моделювання
Наступний патент: Спосіб одержання етанолу з ксилози
Випадковий патент: Спосіб отримання концентратів з цитрусової та плодово-ягідної сировини